

서언
재료 및 방법
조사지 개황
현지조사 방법
통계 및 분석 방법
결과 및 고찰
종면적곡선(species area curves)
환경인자에 따른 토양 화학적 성질
생태적 지위와 분포특성
지표종 분석
적 요
서언
생태적 지위(ecological niche)란 생물간의 상호관계, 환경간의 관계 등을 이유로 형성되는 지위를 뜻한다. 즉, 생물종의 서식처 범위와 먹이사슬 및 경쟁관계에서 차지하는 자원경쟁의 지위를 합한 것을 뜻한다. 생태적 지위를 나타내는 개념은 크게 2가지로 구분된다. ①기본적인 생태적 지위(fundamental niche)와 ②현실 생태적 지위(realized niche)이다. 전자의 경우 한 생물종이 타 생물종에 의하여 자원이나 서식처에 대한 경쟁을 받지 않는 상태를 뜻하며, 후자의 경우 생물종 간의 경쟁에 의한 상태이다(Pulliam, 2000; Silvertown, 2004). 전자의 경우 경쟁이 없으므로 한 생물종이 최대로 수득할 수 있는 자원의 최대치를 의미한다. 한편 지표종(indicator species)은 Müller-dombois and Ellenberg (2003)에 의하여 주로 정의되어 진 개념으로 임업, 농업 그리고 응용생태학에 널리 적용되고 있으며 주로 환경 특성의 생물학적 지표로서 주로 유럽에서 사용되며 생물다양성의 보전으로 주로 이용되는 개념이다(Diekmann, 2003; Simon et al., 2013). 지표종은 다수의 과학자 및 임업관련 전문가들에게 비판을 받아 온 개념이지만, 식물의 생장・생리변화와 개체군간 그리고 환경인자와의 변화추세를 분석하기 위해 널리 이용되어왔다(Diekmann, 2003). 효과적인 지표종의 설정은 ①광범위한 생물다양성의 상태를 반영해야 하며, ②환경적인 상태안에서의 장해요인이 되는 변화를 초기에 알려주는 반응성이 있는 종이어야 한다. 마지막으로 ③예측가능한 범위 내에서 변화에 반응해야 하는 종이다(Simon et al., 2013). 이러한 지표종은 환경구배에 따른 종조성에 영향을 주는 종이다. 이러한 지표종이 소속된 산림환경은 시간에 의한 변화 그리고 기후, 지형조건과 같은 생태적 요소에 의하여 복잡성 및 다른 성질을 갖게 된다(Son et al., 2016). 이에 따라 지형조건에 따른 식물자원의 공간분포양상의 특성을 파악하는 것은 중요하며 향후 이러한 연구의 축적은 효율적인 식물자원확보를 위한 기초연구자료가 된다. 본 연구대상지인 경상북도 청송군에 위치한 면봉산은 보현산 일대에 위치하며 군락 내 인위적인 접근성이 떨어져 외부의 간섭을 적게 받아 임분 내 종조성과 계층구조가 건전하여 생태계의 안정성이 유지되는 곳이다(National Institute of Ecology, 2014). 또한 면봉산 일대 상층부에는 대부분 신갈나무군집이 우점하고 있고, 관목층에는 생강나무, 쇠물푸레 등이 주로 생육하고 있어(Park et al., 2016) 이는 Kim and Lee (2006)가 한반도 삼림식생형에 따른 식물사회학적 식생분류체계에 대한 연구 중, 중‧남부 산지 산림식생형을 신갈나무-생강나무군단으로 분류한 것으로 보아 본 연구대상지는 우리나라 중‧남부지역 온대활엽수림의 종조성과 유사하게 나타나는 곳이다. 본 연구의 목적은 온대중・남부 지역에 속하는 면봉산 일대 식물을 대상으로 지형 및 고도에 따른 식물종의 생태적 지위와 지표종을 관찰하여 식물자원의 공간적 분포패턴의 경향을 파악하고 본 연구대상지 일대의 보전과 관리를 위한 기초자료를 마련하는 것이다.
재료 및 방법
조사지 개황
본 연구대상지는 경상북도 중부지역에 속하며 행정구역상 위치는 경상북도 청송군 현동면 월매리이며 경북대학교 학술림에 속한다(Fig. 1). 서쪽으로 청송군 현서면 무계리, 남쪽으로는 포항시 죽장면 두마리, 동쪽으로는 포항시 죽장면 봉계리와 접하고 있다. 면봉산 일대는 소백산맥의 남단에 위치하며, 주변에 보현산(1,124 m)이 위치하여 상대적으로 해발고도가 높고 지형이 복잡하다. 본 연구대상지는 북위 36°10′42.3″~36°12′17.8″이며, 동경 128°59′45.9″~129°01′21.5″ 사이에 위치한다. 연구대상지를 중심으로 북쪽에 월매저수지가 있고, 지형이 험하고 길고 짧은 계곡이 곳곳에 형성되어 있다. 청송군의 최근 9년간(2010년~2018년) 연평균 기상 관측값을 조사하였다. 연평균기온은 11.1℃, 연평균최고기온 18.1℃, 연평균최저기온 4.9℃, 평균강수량 923.4 ㎜, 평균풍속 1.36 m/s 으로 나타났다. 청송군은 경상북도 내륙지역으로 대륙성기후이며, 여름과 겨울의 기온차가 심하게 일어나고 6월에서 8월 말인 하절기에 강우가 집중되는 경향이다(Korea Meteorological Administration, 2019).
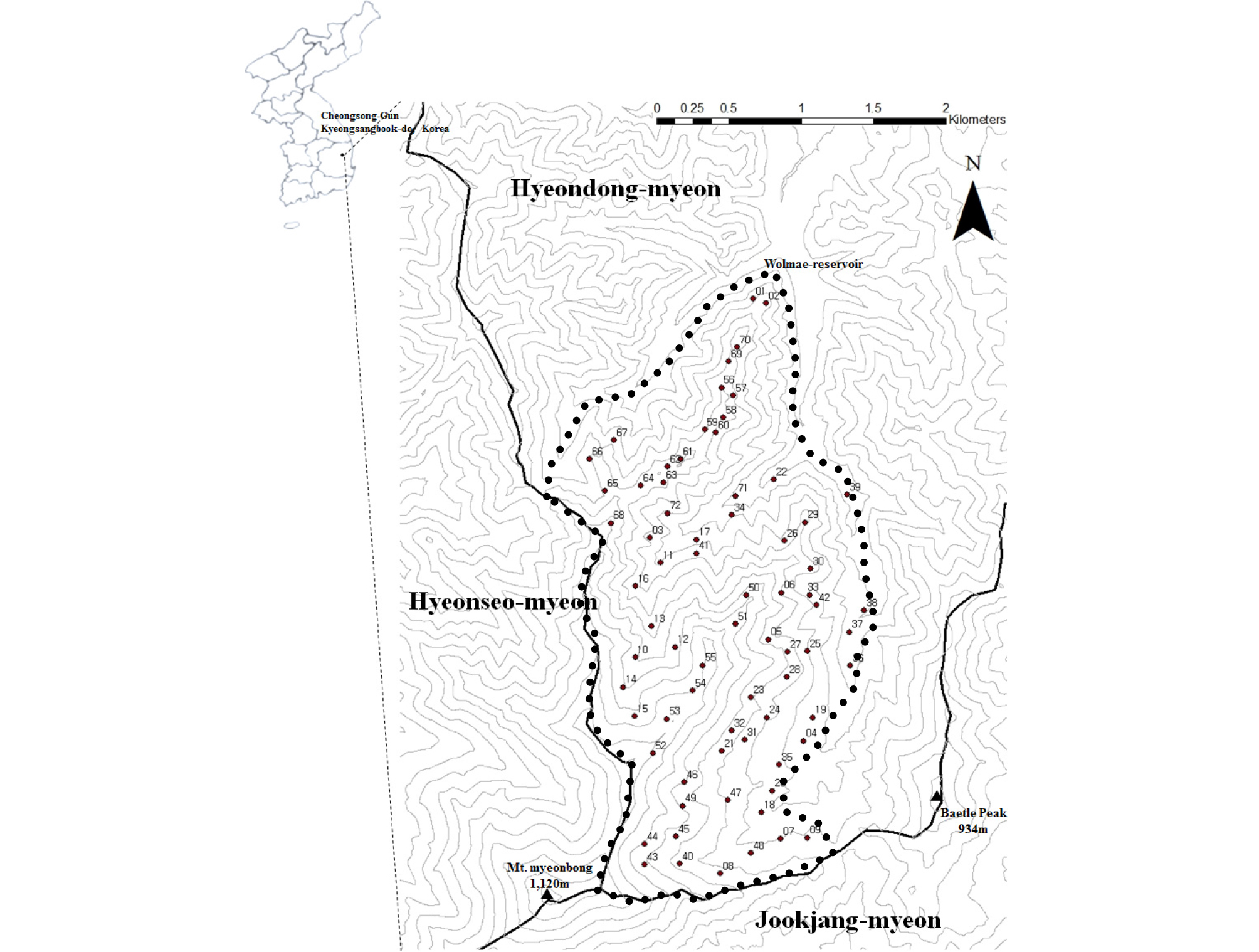
연구대상지 일대는 중생대 백악기 시대에 형성되어 지층은 경상계 산성화산암류, 안산암질암이며, 갈색산림토로 이루어져 있으며 사면부에 위치하고 있다. 토성은 사양토이며 암석노출도는 약 35%로 나타났다(Korea Innovative Solution of Geo- Technology to Sustainable Earth, 2014).
현지조사 방법
본 연구대상지 내 산림식생의 공간적 분포특성에 관한 연구를 위해 방형구의 설치조건은 조림지를 제외하고 가장자리효과(edge effect)가 최소화 되도록 설정하였다. 수치지형도를 참고하여 지형적 특성(산정 및 능선부, 사면부, 계곡부)을 고려하여 총 72개소의 정방형구를 20 m×20 m의 크기로 설치하였다. 방형구 내 흉고직경 4 ㎝ 이상의 모든 입목에 대하여 수종, 흉고직경, 수고를 조사하였으며, 본 연구대상지의 임분구조를 고려하여 상층(upper layer)은 12 m 이상, 중층(middle layer)은 2~12 m 미만으로 지정하였다. 정방형구 내 5 m×5 m의 하층식생(관목층, 초본층) 조사구를 각각 3반복 설치하였다. 관목층 및 초본층 조사과정은 식물종을 동정한 후 Braun-Blanquet (1964)의 식물사회학적 조사방법을 이용하여 피도(coverage)와 군도(dominance)를 측정하였다. 식물의 동정은 원색식물도감(Lee, 2003a; Lee, 2003b)에 의거하였으며, 양치식물의 동정은 한국양치식물도감(Korea Fern Society, 2005)을 이용하였다. 학명과 국명은 국립수목원과 한국식물분류학회가 제시한 국가표준식물목록(Korea National Arboretum, 2019)에 준한 명명을 사용하였다.
통계 및 분석 방법
본 연구대상지 내 적절한 방형구가 설치되었는지에 대한 여부를 판단하기 위해 종면적곡선(Species Area Curves)을 분석하였다(Newton, 2007). 종수 추정을 통한 종면적곡선에서 누적조사구의 순서는 72개 조사구에 일련번호를 정한 후 난수표에 의하여 무작위로 선정하였으며, 수관식생층(상층, 중층)과 하층식생층(관목층, 초본층)으로 나누어 분석하였다. 종풍부도(Number of species, Species richness)에 따른 종수 추정 방법은 Chao 1 (Chao, 1984)의 방법을 이용하였다.
토양의 이화학적 성질을 분석하기 위하여 식생조사가 이루어진 72개의 조사구에서 낙엽층을 걷어낸 후, 5~10 ㎝ 정도의 유기물층을 제거한 곳에 15~20 ㎝의 깊이에서 시료를 채취하였다. 채취한 토양은 밀봉한 상태로 실험실로 운반하여 실내에서 2주간 풍건 후 2 ㎜와 0.02 ㎜의 체에 통과시켜 분석용 시료를 제작하였다. 토성은 비중계법으로 측정하였으며, pH는 풍건토 10 g과 증류수 50 mL를 1:5의 비율로 제작한 후 pH-meter를 사용하여 측정하였다. 유효인산(Available phosphate : Avail. P205)은 Lancaster법을 사용하여 분광분석기(Shimadzu UV-120-02)를 이용하였다. 치환성양이온의 경우 flame photometer (Buck Scientific, PFP7)를 이용하여 측정하였다.
생태적 지위란 서식지 환경에 따라 생육하는 생물종의 최저・최고 내성한계를 나타내는 것으로(Yeocheon Ecological Society, 2005), 특정 환경인자 내에서 생태적 지위폭이 크게 나타나면 환경에 대한 내성의 폭이 넓어 광범위한 지역에서 개체수가 풍부하고 환경에 적응이 잘 되는 종으로 제너럴리스트(generalist)에 가까운 식물종이다. 생태적 지위폭이 낮게 나타나면 환경에 대한 내성이 좁으며 한정적인 지역에서 생육하는 스페셜리스트(specialist)에 해당하는 식물종으로 해석된다. 생태적 지위폭(Ecological niche breadth)의 산정은 Levins(1968)가 제안한 공식을 이용하였으며, 그 공식은 다음과 같다.
Where,
E : Levin’s Ecological niche breadth
Pik: Realative species composition of a given species(i) to the whole gradient that in realized gradient ‘S’
S : Total gradients
수평적 분포에 해당하는 환경구배는 조사지 내에서 조사된 3가지 지형유형(산정 및 능선, 사면, 계곡)으로 선정하였다. 수직적 분포의 경우 800 m 이상, 700 m 이상~800 m 미만, 600 m 이상~700 m 미만, 600 m 미만으로 100 m 간격으로 4가지의 환경구배를 설정하여 각각의 생태적 지위폭을 나타내었다. 72개 조사구 내 발견된 식물종 중에서 빈도수가 3회 이하로 나타난 종은 우연출현종으로 간주하여 생태적 지위의 계산 대상에서 제외하였다. 수평적・수직적 분포역의 평균값을 이용하여 각 층위별 상위 7순위에 해당하는 식물종을 제너럴리스트, 하위 7순위를 스페셜리스트로 지정하였다.
지표종 분석(indicator species analysis)은 미세한 환경의 물리적 환경인자와 토양의 이화학적 성질을 통하여 시행하였다. 지표종 분석은 확률화 검정(randomization test)을 이용한 것으로 현재 특정지역에서 우점하여 현존하는 종으로(Dufrene and Legendre, 1997), 향후 특정 환경인자에 있어 보전과 관리에 핵심이 되는 종으로 해석한다. 통계분석은 PC-ORD (ver. 6.0), SPSS (ver. 19.0)를 이용하였다(McCune and Mefford, 2006).
결과 및 고찰
종면적곡선(species area curves)
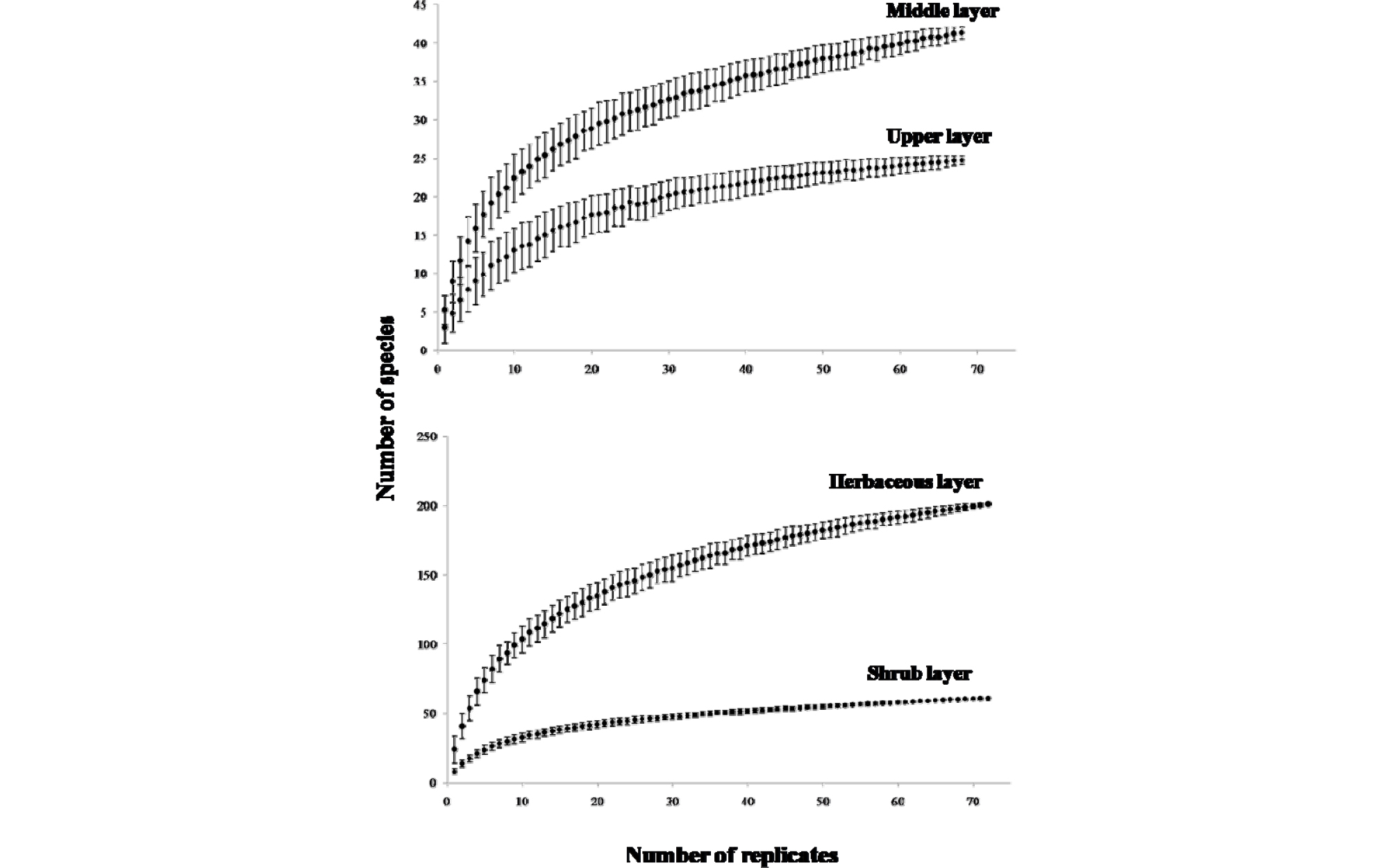
연구대상지 내 적절한 방형구의 설치여부를 알아보기 위하여 수관층식생과 하층식생으로 나누어 각 층위별 종수 추정을 통한 종면적곡선을 분석하였다(Fig. 2). 종수 추정에 있어 조사구 면적수에 따른 종풍부도에 대한 기울기가 0에 수렴하여 본 연구대상지내 식생분석을 위한 적절한 방형구 개소수를 설치하였다고 판단된다.
환경인자에 따른 토양 화학적 성질
본 연구대상지의 환경인자에 다른 토양의 화학적 성질은 Table 1과 같다. 전체 조사구를 기준으로 pH는 4.86, 유기물은 52.53 g/㎏, 유효인산 7.57 ㎎/㎏, 치환성양이온 중에서 칼륨 0.18 cmol+/㎏, 칼슘 2.45 cmol+/㎏ 그리고 마그네슘 0.49 cmol+/㎏으로 분석되었다. 산림토양은 단시간으로 관찰하였을 때는 정지상태로 보이지만 지속적으로 중력방향으로 이동하므로 토양에 유기물을 공급하는 낙엽층, 수분 등이 중력에 의하여 흘러내려갈 확률이 높을 것으로 분석하였다. 해발고도는 유기물층에서 유의한 차이가 나타났는데, 800 m 이상의 그룹에서 61.56 g/㎏으로 가장 높게 나타났다. 이는 700 m 이상 대체적으로 신갈나무의 임분이 두드러지게 나타나(Kim and Kim, 2014) 신갈나무 낙엽에 의한 낙엽층의 형성 및 맹아로 자란 신갈나무 유령목의 고사와 관련 있다고 판단되며 해발고도에 따른 종조성의 차이와 특정 해발고도에서의 바이오매스량의 이입․이출 상관관계에 대한 추가적인 연구가 필요할 것으로 판단된다. 지형의 경우 pH와 칼슘함량에서 유의한 차이가 나타났으며, 계곡부에서 pH가 5.04±0.43으로 가장 높으며 산정부에서 4.59±0.42로 타 지형조건에 비하여 강한 산성토양으로 나타났다. 칼슘을 포함한 모든 토양의 화학적 성질에서 능선 및 산정부의 수치가 낮게 나타났다.
Table 1. Soil chemical properties each environmental factor (±SD, 'a and b' indicates that the item was shown significant different differences among treatments at the 5% level, ANOVA, duncan's post-hoc)
생태적 지위와 분포특성
본 연구대상지 내 조사된 모든 식물종의 수직적 및 수평적 분포역의 상관관계에 대하여 나타낸 결과는 Table 2와 같다. 층위에 따른 각 식물종들의 상관관계 계수와 유의성 판정을 통하여 수직적․수평적 분포역 간에 어떠한 관계가 있는지에 대하여 살펴보았다. 층위별로 두 인자에 대한 상관관계분석을 실시한 결과, 모든 층위의 상관계수는 정의 상관관계로 나타났으나 유의적인 차이는 없는 것으로 분석되었다(coeffecient range: 0.62 ~0.68, p > 0.05). 식물은 환경에 따라 생존을 위한 제각기 다른 생리활성을 가지고 생육한다(Salisbury and Ross, 1992). 생존에 필요로 하는 수분, 광, 온도 등 환경의 변화로 인한 내성범위와 분포의 제한요인은 각 식물이 가진 생리적 매커니즘에 따라 다르다. 이러한 환경변화 자극에 대한 반응을 통해 식물은 이에 적응하게 되므로 생태적 지위 또한 시간의 흐름에 따라 변화하는 것으로 판단된다.
Table 2. Correlation coefficient between horizontal and vertical ecological niche breadth (Pearson's Correlation method was used)
Layer Index | Upper (n=25) | Middle (n=42) | Shrub (n=61) | Herbaceous (n=201) |
| Correlation coefficient | 0.68 | 0.63 | 0.65 | 0.62 |
| p-value | 0.08 | 0.34 | 0.30 | 0.40 |
수관층 생태적 지위
수관층 및 하층식생에 속하는 식물의 수평적 및 수직적 분포역을 살펴본 결과 Table 3, 4와 같다.
Table 3. Gerneralist and specialist species through Ecological niche breadth of crown vegetation in study sites (Rank for 7 species for each part, G: Generalist, S: Speciealist)
Table 4. Gerneralist and specialist species through Ecological niche breadth of understory vegetation in study sites (Rank for 7 species for each part, G: Generalist, S: Specialist)
상층(Upper layer)에서 생태적 지위를 기반으로 한 제너럴리스트는 신갈나무, 소나무, 굴참나무, 고로쇠나무, 물푸레나무, 쪽동백나무, 박달나무로 나타났으며, 수평적 분포역은 0.571 ~0.799, 수직적 분포역은 0.644~0.942로 나타났으며, 그 평균값은 0.607~0.871로 나타났다. 수평적 분포역이 가장 높은 수종은 소나무이며, 수직적 분포역이 가장 높은 수종은 신갈나무로 분석되었다. 소나무는 교목성 침엽수로 수평적, 수직적 분포역이 넓고 수직적 분포의 경우에도 우리나라에서 100~1,300 m 일대에 분포하고 있다(Kong, 2004). 신갈나무는 설악산에서 해발고도 200~1,700 m, 지리산에서는 600~1,800 m의 수직적 분포역을 가지며, 백두산, 설악산 지리산, 한라산 등에 걸쳐 넓은 수평적 분포역을 가지는 수종이다(Kong et al., 2014). 제너럴리스트에 해당하는 고로쇠나무와 물푸레나무의 경우 수평적 분포역이 수직적 분포역에 비하여 현저히 적은수치로 나타나 타 수종과 비교하여 고도에 대한 분포영향보다 지형조건과 같은 수평적 분포역 영향에 민감하게 반응하는 것으로 판단된다. 고로쇠나무는 내한성이 강하고 음수성이 강한 수종으로(Cho, 1990) 온도의 내성범위가 넓은 수종으로, 수직적 분포역이 수평적 분포역에 비해 상대적으로 높게 나타난 결과라고 판단된다. 스페셜리스트는 굴피나무, 서어나무, 떡갈나무, 당단풍나무, 산뽕나무, 피나무, 까치박달로 선정되었으며, 수평적 분포역은 모두 0.333으로 낮은 수치를 기록하였다. 수직적 분포역은 0.622~0.359로 나타났다. 까치박달 또한 고로쇠나무와 유사하게 비교적 내한성이 강한 수종에 속하여(Korea National Arboretum, 2019) 수직적 분포역은 수평적 분포역에 비하여 높은 수치를 나타내었다. 본 연구대상지 내에서 상층부에서 가장 낮은 생태적 지위로 분석된 수종은 굴피나무로 나타났다. 본 연구대상지 일대에서는 임도주변 및 계곡주변의 양지바른 곳에서 군락형태보다는 단목형태로 생육하고 있었다.
중층(Middle layer)에서 제너럴리스트에 선정된 식물은 박달나무, 신갈나무, 팥배나무, 피나무, 쇠물푸레나무, 굴참나무, 당단풍나무로 조사되었다. 신갈나무는 수관층식생에서 상층과 중층의 수직적․수평적 분포를 우점하고 있는 식물종이다. 기 연구된 자료에 따라, 면봉산 일대의 주요 우점군집을 신갈나무와 신갈나무-소나무 군집으로 명명한 것과 유사한 연구결과로 나타났다(Park et al., 2016). 생태적 지위폭은 수평적 분포역 0.582~0.857, 수직적 분포역 0.694~0.940으로 나타났다, 당단풍나무의 경우 수평적 분포가 0.582, 수직적 분포가 0.919로 나타났다. 단풍나무속(Acer spp.)에 속하는 수종의 경우, 고도변화에 따른 온도차에 대한 내성범위가 높아(Cho, 1990). 상층에서 고로쇠나무의 수직적 분포역이 수평적 분포역보다 높은 수치를 나타내는 것으로 보아 상응하는 연구결과로 분석되었다. 스페셜리스트로 선정된 수종은 비목나무, 졸참나무, 고추나무, 진달래, 참회나무, 물오리나무, 서어나무로 나타났다. 서어나무의 경우 우리나라의 극상림(climax)를 대표하는 수종중 하나이다(Son et al., 2016). 하지만 본 연구대상지 내 신갈나무와 소나무의 생태적 지위 분포역이 넓고 서어나무는 좁은 것으로 보아 아직까지 극상림에 도달하기 전의 침활혼효림으로 현재 본 연구대상지의 임분은 수관경쟁기와 하층재형성기에 접어들어 활발한 임분구조 내 종간경쟁이 진행되고 있는 것으로 판단된다(Smith, 1986). 주로 수관밀도가 높으며 공중습도가 높은 계곡부에 한정적으로 생육하고 있었다.
하층식생 생태적 지위
관목층의 제너럴리스트 식물종의 수평적 분포역은 0.714 ~0.920이며, 수직적 분포역은 0.742~0.992로 나타났으며, 주요 식물종은 생강나무, 진달래, 팥배나무, 철쭉, 조록싸리, 함박꽃나무, 개옻나무로 선정되었다. 스페셜리스트 식물종은 산조팝나무, 호랑버들, 굴피나무, 누리장나무, 물참대, 소나무, 산뽕나무로 수평적 분포역이 0.333~0.667이며, 수직적 분포역은 0.250~0.689로 나타났다. 생강나무는 우리나라 신갈나무림에서 대표적인 관목층을 형성하는 수종으로(Kim and Lee, 2006) 본 연구대상지에도 이에 상응하는 연구결과로 나타났다. 스페셜리스트에 속하는 산조팝나무의 경우 능선부의 바위가 많은 지형에서 주로 생육하였으며, 상층부의 소나무임분에서 주로 발견되었다. 누리장나무의 경우 수평적 분포역이 수직적 분포역에 비하여 월등히 높은 수치로 나타났는데 주로 임연부 및 사면하부의 햇빛이 잘 드는 곳에서 자주 발견되었고 수평적 분포역에서 0.667의 수치로 나타났지만 주로 400~500 m의 낮은 해발고도에 한정적으로 분포하여 수직적 분포역에서 낮은 수치를 기록하였다.
초본층의 제너럴리스트는 개옻나무, 조록싸리, 둥굴레, 미역줄나무, 애기나리, 신갈나무, 그늘사초의 순으로 나타났으며, 수평적 분포역은 0.939~0.986, 수직적 분포역은 0.852~0.970으로 계산되었다. 스페셜리스트는 산달래, 노린재나무, 꿩의바람꽃, 오리방풀, 애기괭이눈, 넉줄고사리, 옥잠난초의 순으로 나타났다. 수평적 분포역은 모두 0.333으로 나타났으며, 수직적분포역은 0.361~0.440의 분포로 나타났다. 초본층의 주된 제너럴리스트에 해당하는 식물종 중에서 개옻나무나 싸리류(조록싸리) 그리고 만경류(미역줄나무)와 같은 식물종은 2차림형태의 임분이 재형성되어 가는 시기에 주로 선구수종으로 생육하는 종이다. 이는 임분이 안정화시기에 도달하기 전 수관층에서의 각 개체목간의 치열한 경쟁관계에 따른 자연적 교란과 그에 따른 수관닫힘이 빈번히 발생하여 나타난 결과로 판단된다. 이러한 천이 초기 구성종은 상층부의 수관층이 발달되고 성숙한 임분으로 진행됨에 따라 향후 하층구성종의 변화를 살펴보기 위한 장기적인 모니터링이 요구된다. 스페셜리스트에 해당하는 식물종은 대부분 반그늘 또는 그늘진 음지와 계곡부에서 주로 발견되었다. 이러한 종들은 타 식물종과 생육하는 환경의 상관관계에 있어 광경쟁 뿐만 아니라 수분스트레스에 대하여 민감한 종으로 판단된다. 초본층에서 스페셜리스트로 선정된 초본식물 중 옥잠난초, 오리방풀, 꿩의바람꽃, 산달래의 경우 근경이 짧게 분지하는 생육형을 가진다. 이러한 생육형은 매우 좁은 범위의 연결체를 형성하여 생육지 내에서 개체군의 존속을 위하여 최소한의 공간범위와 필요한 자원을 이용하는 생존전략을 가지는 것으로 판단된다
지표종 분석
산림 내 특정 환경 및 조건에서 대표로 서식하는 식물종을 알아보기 위하여 종조성을 기반으로 한 지표종 분석을 실시하였으며, 각각의 환경인자에 따른 그룹별 종조성의 이질성을 파악하기 위하여 MRPP-test (Multi Response Permutation Procedure)를 실시하여 검증하였다. MRPP-test는 각 그룹간 종조성의 차이를 판별하여 해당 그룹간의 유의성 및 유사계수를 이용하여 군집의 이질성을 판단한다(McCune and Grace, 2002).
해발고도에 따른 지표종
해발고도에 따른 지표종을 살펴보기 위하여 800 m 이상, 700 m 이상~800 m 미만, 600 m 이상~700 m 미만, 600 m 미만으로 100 m 간격으로 4가지의 그룹으로 분류하여 분석하였다. MRPP- test 결과(Table 5), 상층과 중층은 주로 해발고도 800 m 이상에 서식하는 종조성과 타 해발고도간의 종조성에 유의적인 차이가 있는 것으로 나타났고, 관목층과 초본층의 경우 600 m 미만의 낮은 해발고도 그룹과 타 해발고도간의 종조성 차이가 있는 것으로 분석되었다.
Table 5. Results of MRPP-test for each altitude group (T: MRPP-test statistic, A: index of similarity within groups)
해발고도에 따른 지표종 분석 결과는 Table 6과 같다. 상층에서는 600 m 미만의 그룹에서만 굴피나무(31.9)와 졸참나무(25.2)로 나타났다. 중층은 800 m 이상에서 철쭉(34.5)으로 나타났으며, 600 m 미만에는 쪽동백나무(52.3), 물푸레나무(39.5), 잔털벚나무(29.1), 서어나무(26.5), 졸참나무(23.9), 생강나무(20.6), 비목나무(17.1)로 나타났다.
Table 6. Indicator species analysis of altitude
관목층은 해발고도 800 m 이상에서 잣나무(16.4)가 지표종으로 선정되었다. 본 연구대상지에서 발견되는 잣나무의 갱신형태는 주로 설치류에 의한 종자운반으로 인하여 발아한 것이 많을 것으로 판단되나 정확한 분석을 위하여 향후 잣나무발생과 동물간의 상호작용에 대한 추가적인 조사가 요구된다. 초본층에서는 해발고도 800 m 이상의 그룹에서 큰참나물(43.3), 십자고사리(39.5), 뫼제비꽃(32.2), 미역취(21.2), 딱총나무(18.8)로 나타났으며, 600 m 미만의 해발고도에서 얼레지(37.5), 비목나무(34.2), 참개암나무(27.3), 졸참나무(25.5), 큰꼭두서니(24.6), 우산나물(20.3), 신나무(18.2), 노린재나무(15.4), 청미래덩굴(14.5), 으아리(13.6)로 나타났다.
졸참나무는 우리나라에 자생하는 참나무류 목본식물 중에서 중부이남에 주로 생육하고 있으며 타 참나무속의 식물에 비하여 낮은 해발고도에서 생육하는 식물종으로(Kim and Kim, 2014), 본 연구결과에서도 상층부와 초본층에서 600 m 미만의 그룹을 대표하는 지표종으로 나타났다. 잣나무는 우리나라의 지리산 이북에 위치하며 높은 산지의 능선부에 위치하고 있으며(Korea National Arboretum, 2019), 본 연구결과에서도 관목층에서 800 m 이상의 높은 고도에서 지표종으로 분석되었다.
본 연구결과로 나온 해발고도에 따른 지표종 분석에서는 해발고도 800 m 이상 그룹과, 해발고도 600 m 미만의 그룹에서만 지표종이 나타났는데, 이는 산림식생의 수직적 분포양상에 있어 종조성의 변화가 약 200~300 m 해발고도 차이에 따라 나타나는 결과라고 판단된다. 해발고도에 따른 식물의 종다양성 패턴은 동일한 산지 내 인접한 조사구간에서 다를 수 있을 정도로 우리나라 산림생태계의 생물종다양성은 복잡하지만 이를 제한하는 인자(서식지조건, 기후, 해발고도 등)는 같을 수 있다(Lee and Chun, 2018). 따라서 우리나라 산지와 같이 지형이 험난한 서식지 조건의 산림생태계에서 종조성에 따른 지표종 또한 다르게 분석될 수도 있다. 우리나라에 분포하는 여러 식생대 및 특이서식지 등에 대한 지표종 분석사례를 종합하여 우리나라 산림생태계를 이해하기 위한 다각적인 분석이 필요하다.
지형에 따른 지표종
지형그룹은 조사구 내에서 현지조사된 지형조건 및 계곡부 유무에 따라 3가시 지형유형(산정 및 능선, 사면, 계곡)으로 선정하였다.
MRPP-test 결과(Table 7) 대체적으로 지형에 따라 모든 층위에서 종조성이 이질적인 그룹으로 유의적인 차이가 있는 것으로 나타났다(p < 0.01).
Table 7. Results of MRPP-test for each landform group (T: MRPP-test statistic, A: index of similarity within groups)
지형조건에 따른 층위별 지표종(Table 8)은 계곡부의 초본층에서 많은 지표종이 나타난 것으로 분석되었다. 상층에서는 산정 및 능선부의 소나무(44.6)로 단일 수종으로 나타났으며 사면부는 신갈나무(44.5), 박달나무(23.0)로 조사되었다, 계곡부는 물푸레나무(37.9), 잔털벚나무(32.2), 졸참나무(27.4), 굴피나무(17.9), 까치박달(17.9), 물박달나무(15.6), 당단풍나무(14.3), 서어나무(14.3)로 나타났다. 중층은 산정 및 능선에서 쇠물푸레나무(48.2), 굴참나무(33.2)로 나타났으며, 계곡부에서 당단풍나무(43.4), 물푸레나무(38.2), 까치박달(28.6), 산뽕나무(27.2), 고로쇠나무(25.8), 쪽동백나무(25.3), 층층나무(25.2), 서어나무(14.3)로 나타났다. 관목층의 지표종 분석결과는 계곡부에서만 분석되었으며, 고추나무(21.4), 참회나무(16.7)로 나타났다. 초본층의 경우 가장 많은 지표종이 나타났는데, 산정 및 능선부에서 싸리(32.9), 기름나물(24.7), 가는잎그늘사초(15.5)로 나타났다. 사면부에서는 노루발(35.7)로 나타났으며, 계곡부의 경우 까치박달(50.0), 얼레지(48.5), 단풍마(37.8), 투구꽃(36.2), 고추나무(35.9), 미나리냉이(35.7), 노루오줌(30.2), 하늘말나리(28.8), 참개암나무(28.6), 얇은잎고광나무(28.4), 개별꽃(27.7), 작살나무(25.2), 큰개별꽃(24.1), 천남성(22.3), 족도리풀(21.0), 비목나무(19.9), 산뽕나무(17.9), 싸리냉이(17.9), 자주잎제비꽃(17.9), 큰괭이밥(17.9), 큰꼭두서니(16.5), 관중(14.3), 나도물통이(14.3), 산달래(14.3), 삿갓나물(14.3), 현호색(14.3)으로 나타났다.
Table 8. Indicator species analysis of landforms (PR: peak & ridge, S: slope, V: valley)
지형조건은 미세환경(광량, 수분, 토양조건 등)의 대동소이한 차이가 나타나며 이에 따른 임분구조의 구성의 변화가 생긴다. 이에 상층임관의 변화로 하층식생의 종구성에 큰 영향을 미친다(Chen and Franklin, 1997; Leith and Aston, 1961). 계곡부는 식물들이 생육할 수 있는 다양한 자원이 충족되어 여러 개체가 군집을 이루며 정착할 수 있는 조건이 마련된 것으로 판단된다(Byun et al., 2013). 계곡부는 다른 지형조건에 비하여 다수의 지표종이 나타난 것으로 분석되었는데 이러한 결과로 보아 계곡부는 다양한 식물종의 정착과 생육에 유효한 기회를 제공하며 개체군 또는 개체간 동적인 생존경쟁이 일어나는 특별한 생태계이다(Müller-Dombois and Ellenberg, 2003).
적 요
본 연구의 목적은 온대중·남부 지역에 속하는 면봉산 일대 식물을 대상으로 지형조건 및 고도에 따른 식물종의 생태적 지위와 지표종을 관찰하여 식물자원의 환경적 구배에 따른 공간적 분포경향을 파악하는 것이다. 수평·수직 생태적 지위의 경우 상층과 중층에서 신갈나무가 높은 생태적 지위를 가지고 있었으며, 상층에서는 수직적 생태적 지위가, 중층에서는 수평적 생태적 지위가 높아 본 연구대상지 일대의 제너럴리스트인 것으로 분석되었다. 또한 관목층에서 생강나무가 제너럴리스트로 분석되어, 한반도 중·남부 산지의 대표식생을 신갈나무-생강나무 군단으로 분류한 것과 유사한 결과를 도출하였다(Kim and Lee, 2006). 지표종 분석 결과, 고도와 지형조건에 따른 환경구배의 차이에서 주로 하층식생이 수관층식생보다 더 이질적인 종조성으로 분석되었다.
