

서언
재료 및 방법
실험재료 및 추출물 조제
세포배양 및 MTT assay
DNA fragmentation 분석
DAPI staining에 의한 세포핵의 형태 관찰
Flow cytometry 분석
Western blot analysis에 의한 단백질 발현 분석
Mitochondrial membrane potential (MMP, Δψm) 변화의 측정
통계 처리
결과 및 고찰
인체백혈병 U937 세포의 증식과 apoptosis에 미치는 ELM의 영향
외인적 경로 Death receptor 및 ligand 발현에 미치는 ELM의 영향
내인적 경로 Bcl-2 family의 발현에 미치는 ELM의 영향
MMP (Δψm) 변화에 미치는 pachymic acid의 영향
IAP family의 발현에 미치는 SSeo의 영향
Caspases의 활성에 미치는 ELM의 영향
적요
서언
인체에서 발병하는 여러 종류의 암 중에서 여섯 번째로 높은 발병률을 가지는 것으로 알려져 있는 혈구암은 백혈구의 비정상적인 증식을 유발시키는 특징을 가지며 혈액 또는 골수에서 유발되는 암이다(Sachdeva et al., 2019). 혈구암은 골수에 위치하는 림프구에서 유발되는 림프구성 백혈병(lymphocytic leukemia)과 과립성 백혈구 및 단핵 백혈구에서 유발되는 골수성 백혈병(myelogenous leukemia)으로 구분되어 진다(Acheampong et al., 2018; Gilliland et al., 2004). 혈구암은 항암치료에 대해서 대부분 저항성을 가지고 있어 치료가 어려우며 재발할 확률도 매우 높은 것으로 보고되어 있으며, 혈구암의 치료에 대한 다양한 종류의 항암제들은 개인의 특성에 따라 약리작용과 독성에 의한 부작용이 문제점으로 지적되고 있다. 따라서 혈구암을 치료에 보다 효과적인 생리활성을 갖는 소재를 발굴하고 그와 관련된 분자 및 세포수준에서의 기전을 밝히는 것이 중요하다고 할 수 있다.
암의 효과적인 예방과 치료에 있어서 세포사멸(apoptosis) 과정을 통하여 비정상적인 세포나 암세포를 제거하는 치료제의 개발이 최근 중요시되고 있으며, 이러한 apoptosis는 정상세포나 주위의 조직에 손상을 주지 않으면서 악성 또는 암세포 등의 제거를 위한 생리적 기전으로 세포의 위축(cell shrinkage)과 염색질 응축(chromatin condensation)과 같은 형태적 변형이 동반되는 세포죽음의 일종이다. Apoptosis 경로는 내인적 경로(intrinsic pathway) 및 외인적 경로(extrinsic pathway)로 구별되어진다. 내인적 경로는 미토콘드리아 막의 붕괴에 따른 세포질로의 cytochrome c와 같은 인자의 방출을 통한 caspase-9의 활성을 통하여 apoptosis를 유도한다(Jan and Chaudhry, 2019; Tay et al., 2019). 외인적 경로는 사멸수용체(DRs)와 ligand와의 상호작용을 통한 caspase-8의 활성에 의하여 이루어진다(Ivanisenko and Lavrik, 2019). 또한 활성화된 caspase-8은 세포질에 존재하는 Bid를 절단형(truncated Bid, tBid)으로 전환시켜 pro-apoptotic 단백질인 Bax의 중합화(oligomerization)에 의한 미토콘드리아 삼투성 전이 미세공(mitochondrial permeability transition pore)의 형성을 통하여 mitochondrial membrane potential (MMP)의 소실과 cytochrome c의 세포질로의 유리를 유도한다(Han et al., 2008; Lazebnik et al., 1994; Lee et al., 2010). Apoptosis의 최종적 조절인자인 caspase-3 및 -7과 같은 실행자 caspase가 활성화되어 표적 기질 단백질들의 분해를 통하여 apoptosis를 유도시키며, caspase 효소들은 inhibitor of apoptosis proteins (IAPs) family에 속하는 단백질들과의 직접적인 결합을 통하여 그들의 apoptotic 활성을 억제시킨다.
부처꽃(Lythrum anceps (Koehne) Makino)은 쌍떡잎식물강-도금양목-부처꽃과의 여러해살이풀로 냇가, 초원 등의 습지에서 자라며, 주로 한국, 일본, 유럽 등지에서 분포한다. 예로부터 전초를 청혈(淸血), 지혈(止血), 양혈(凉血)의 효능으로 이질, 방광염, 이뇨, 지사제(止瀉劑) 등을 치료하기 위해 전통적으로 널리 사용되어 왔다. 부처꽃의 주요 생리활성 성분은 전초에는 salicarin, tannin이 함유되어 있고, 꽃에는 vitexin, orientin, malvin, cyanidin-3-monogalactoside, gallic acid, ellagic acid, 및 소량의 chlorogenic acid가 함유되어 있다(Piwowarski et al., 2015). 부처꽃의 효능에 대한 연구보고를 살펴보면, 꽃잎에서 조제된 추출물의 DPPH 및 리놀레산에 대한 항산화 효능에 대한 보고와 실험동물에서의 항염증 및 항통증 효과(Tunalier et al., 2007)가 보고되었고, 과혈당 마우스에서의 효과(Lamela et al., 1986) 및 정상 혈당 흰쥐에 대한 효과(Lamela et al., 1985), 항미생물 효과(Becker et al., 2005) 및 항미생물 화합물(Rauha, 2001)에 대한 보고가 되어있으며, 국내에서는 부처꽃 뿌리 추출물의 항산화 및 간장 보호활성(Lee et al., 2009a), 사염화탄소로 섬유화가 유도된 흰쥐 간에서 부처꽃 뿌리 추출물의 항산화 및 섬유화 저해 활성 (Lee et al., 2009b)에 관해 보고되었다. 하지만 부처꽃 추출물에 의해 유도되는 apoptosis 과정과 그에 따른 분자생물학적 기전에 대해서는 명확히 밝혀져 있지 않다.
따라서 본 연구에서는 부처꽃 에탄올추출물이 인체백혈병 U937 세포에 미치는 항암 효과의 생화학적 기전의 해석을 위하여 U937 세포의 증식에 미치는 영향을 조사하였고, 부처꽃 에탄올 추출물이 유발하는 증식억제 현상이 apoptosis에 어떠한 영향을 미치는지를 조사하여 유의적인 결과를 얻었기에 이를 보고하는 바이다.
재료 및 방법
실험재료 및 추출물 조제
본 실험에 사용한 부처꽃 시료는 경북 예천군 풍양면 효갈리(2017년 7월)에서 인식형질이 있는 개체를 직접 채집하여 건조 파쇄 하였으며, 본 표본은 국립낙동강생물자원관 수장고에 보관하고 있다(NNIBRVP60168). 식물의 동정은 ‘Korea Biodiversity Information System’ 의거하여 분류를 실시하였다(국가생물종지식정보시스템-Korea Biodiversity Information System, 2014). 부처꽃 전초 에탄올 추출물을 얻기 위하여 건조 시료 무게의 30배에 해당하는 70% 에탄올을 제조하여 1시간 초음파 추출 후 실온에서 24시간 동안 방치하였다. 에탄올 추출물을 여과지(Whatman No. 3 filter paper, Whatman International Ltd., Maidstone, England)로 여과한 후 용매를 Rotary evaporator (Tokyo Rikakikai Co., Tokyo, Japan)를 이용하여 제거하였고, 동결건조(Ilshin, Korea)를 거쳐 건조중량 대비 9.6%의 분말 시료를 얻었다. 이후 에탄올 추출시료는 dimethyl sulfoxide에 20 ㎎/mL의 농도로 녹여 –20℃에서 보관하였으며, 실험에 따라 적절하게 희석하여 사용하였다.
세포배양 및 MTT assay
인체 급성 골수 백혈병(acute myeloid leukemia) U937 세포는 American Type Culture Collection (Manassas, VA, USA)에서 구입하였으며, 10% fetal bovine serum (FBS)가 함유된 RPMI 1640 배지(Rockville, MD, USA)를 이용하여 37℃, 5% CO2 조건에서 배양하였다. ELM 처리에 따른 세포생존율 정도를 측정하기 위하여 6-well plate에 U937 세포를 well당 3×105개를 분주하고 ELM을 적정 농도로 처리하여 배양하였다. 이후 MTT 시약을 0.5 ㎎/mL 농도가 되게 처리하여 2시간 동안 배양하고, 배지를 제거한 후 DMSO를 2 mL씩 분주하여 생성된 formazan을 모두 녹인 후 ELISA reader (Molecular Devices, CA, USA)로 540 ㎚에서 흡광도를 측정하였다.
DNA fragmentation 분석
Apoptosis 유발의 또 다른 증거를 제시하기 위하여 DNA 단편화 현상의 분석을 실시하였다. 상기와 동일한 방법으로 처리된 세포들을 모은 다음 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100]를 첨가하여 상온에서 1시간 동안 처리한 다음 상층액을 회수하였다. 회수된 상층액에 proteinase K solution (Sigma-Aldrich, St, Louis, MO, USA)을 처리하여 50℃에서 3시간 동안 반응시킨 후 phenol : chloroform : isoamyl alcohol (25:24:1) 혼합 용액(Sigma- Aldrich, St, Louis, MO, USA)을 첨가하고 30분간 회전교반 시킨 다음 원심분리하여 다시 상층액을 분리하였다. 분리된 상층액에 isopropanol (Sigma-Aldrich, St, Louis, MO, USA)과 5 M NaCl를 첨가하여 4℃에서 24시간 동안 반응시킨 후 원심분리시켜 DNA pellet을 추출하였다. 추출된 DNA pellet에 RNase A가 포함된 TE buffer 및 6X gel loading dye (Bioneer, Daejeon, Korea)를 첨가하고 1.6% agarose gel을 이용하여 50 V 전기영동 시킨 후 ethidium bromide (Sigma-Aldrich, St, Louis, MO, USA))로 염색하여 DNA 단편화 현상을 확인하였다.
DAPI staining에 의한 세포핵의 형태 관찰
Apoptosis가 유발되었을 경우 특이적으로 나타나는 핵의 형태적 변화를 관찰하기 위하여 LEM이 처리된 세포를 모은 다음 37% formaldehyde 용액과 PBS를 1:9의 비율로 섞은 fixing solution으로 10분 동안 고정하였다. 고정된 세포를 cytospin을 이용하여 slide glass에 부착하였다. 세포가 부착된 slide glass를 PBS로 2~3회 정도 세척하고 PBS가 마르기 전에 0.2%의 Triton X-100 (Amresco)을 첨가하여 상온에서 10분간 고정한 후 2.5 ㎍/mL 농도의 4',6-diamidino-2-phenylindole (Sigma- Aldrich, St, Louis, MO, USA) 용액을 처리하여 상온에서 15분간 염색하였다. 염색이 끝난 후 400배의 배율로 각 농도에 따른 암세포의 핵의 형태 변화를 관찰하였다.
Flow cytometry 분석
U937 세포에서 ELM이 유발하는 apoptosis 정도를 정량적으로 분석하기 위하여 정상 및 ELM이 24시간 동안 처리된 세포들을 모은 다음 2,000 rpm으로 5분간 원심분리하여 상층액을 제거한 후 PBS를 이용하여 2~3회 정도 세척하였다. 준비된 세포는 CycleTEST PLUS DNA REAGENT Kit (Becton Dickinson, San Jose, CA, USA)를 이용하여 고정 및 염색을 하여 4℃, 암실에서 30분 동안 반응을 시켰다. 반응시킨 세포를 35 ㎜ mesh를 이용하여 단일세포로 분리한 후 FACSCalibur (Becton Dickinson)를 적용시켜 형광반응에 따른 cellular DNA content 및 histogram을 CellQuest software 및 ModiFit LT (Becton Dickinson) 프로그램을 이용하여 분석하였다.
Western blot analysis에 의한 단백질 발현 분석
준비된 세포에 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가하여 총 단백질을 분리하였다. 동량의 sample을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리한 후, nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electro-blotting에 의해 전이시켰다. 분리된 단백질이 전이된 membrane을 5% skim milk를 처리하여 비특이적인 단백질들에 대한 blocking을 실시하고 1차 항체를 처리하여 상온에서 2시간 이상 또는 4℃에서 over night 시킨 다음 1차 항체에 맞는 2차 항체를 사용하여 상온에서 1시간 정도 반응시켰다. 반응이 끝난 후 enhanced chemiluminoesence (ECL) solution (Amersham Life Science Corp., Arlington Heights, IL, USA)을 적용시킨 다음 X-ray film에 감광시켜 특정단백질의 발현 양을 분석하였다. 본 연구에 사용된 항체는 Santa Cruz Biotechnology (Santa Cruz, CA, USA), Chemicon (Temecula, CA, USA) 및 Amersham Life Science Corp.에서 구입하였다.
Mitochondrial membrane potential (MMP, Δψm) 변화의 측정
MMP 변화 정도를 측정하기 위하여 적정시간 동안 ELM이 처리된 세포들을 모은 다음 5,5',6,6'-tetrachloro-1,1',3,3'- tetraethyl-imidacarbocyanine iodide (JC-1, Sigma-Aldrich, St, Louis, MO, USA) 용액을 처리하여 20분 동안 37℃에서 반응시켰다. 반응시킨 세포를 2,000 rpm으로 원심 분리하여 상층액을 버리고 다시 500 μL의 차가운 PBS를 첨가하고 35 ㎜ mesh를 이용하여 단일세포로 분리한 후 유세포 분석기에 적용시켜 ELM 처리에 따른 MMP의 변화 정도를 분석하였다.
통계 처리
실험 결과들의 유의성을 검정하기 위하여 분산분석(ANOVA)을 실시한 후 p<0.05 수준에서 Duncan’s multiple range tests를 실시하였으며, 그 결과는 평균(mean) ± 표준편차(standard deviation, SD)로 표시하였다. 이때 사용한 모든 통계분석은 SPSS 17.0 (IBM SPSS Inc, NY, USA) 통계 프로그램을 이용하여 처리하였다.
결과 및 고찰
인체백혈병 U937 세포의 증식과 apoptosis에 미치는 ELM의 영향
인체백혈병 U937 세포에서 ELM이 유발하는 세포증식 억제 현상을 조사하기 위하여 ELM을 24시간동안 적정농도로 처리한 후 MTT assay를 실시한 결과, 25 ㎍/mL 이상 처리군에서부터 ELM 농도 의존적으로 생존률 및 증식억제 효과가 나타났다(Fig. 1A). ELM 처리에 의한 U937 세포의 증식억제가 apoptosis 유도와 연관성이 있는지를 조사하기 위하여 apoptosis 유발 시 특이하게 관찰되는 핵의 형태변화와 어떠한 연관이 있는지를 DAPI 염색법으로 확인하였다. Fig. 1B에 나타낸 바와 같이 정상에서 배양된 암세포와는 다르게 ELM을 처리한 배지에서 배양된 세포에서 처리농도 의존적으로 핵의 밀도 감소와 더불어 apoptosis 과정 중 전형적으로 관찰되는 염색질 응축에 의한 apoptotic body가 관찰되었다. 다음으로 apoptosis 유발의 또 다른 증거인 DNA 단편화 현상을 관찰한 결과 Fig. 1C에 나타난 바와 같이 ELM 처리 농도가 증가할수록 DNA 단편화 현상이 관찰되었다. ELM 처리에 의한 apoptosis 유발의 정량적인 평가를 위하여 동일한 조건으로 배양된 U937 세포를 대상으로 propidium iodide (PI) 염색 후 DNA flow cytometry를 이용하여 sub-G1기에 해당하는 세포주기 분포도를 조사한 결과, Fig. 1D에 나타낸 바와 같다. 정상배지에서 배양된 암세포에서 자연적 apoptosis 유발 빈도는 약 1.24%로 매우 낮았으나, ELM 처리농도 증가에 따라 sub-G1기에 해당하는 세포의 빈도가 증가하여 25, 50 및 100 ㎍/mL 처리군에서 약 5.76%, 28.32% 및 54.68% 정도의 apoptosis가 처리농도 의존적으로 증가하였음을 알 수 있었다.
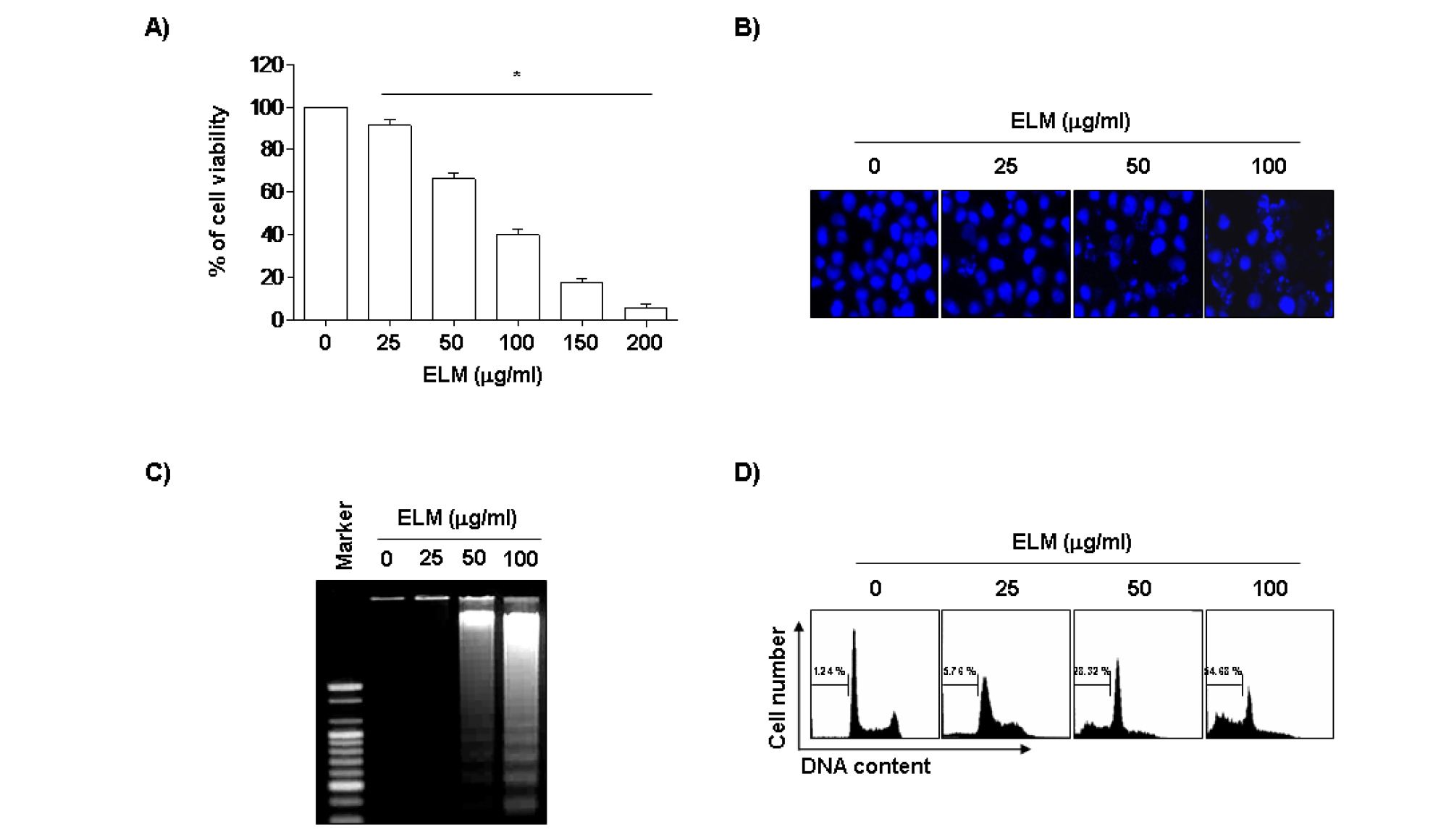
Fig. 1.
Inhibition of cell viability and induction of apoptosis by ELM treatment in human leukemia U937 cells. Cells were 6-well plated at 3×105 cells, and incubated for 24 hr. The cells were treated with the indicated concentrations of ELM for 24 hr. (A) Cell viability was measured by the metabolic-dye-based MTT assay. (B) Cells grown under the same conditions as (A) were fixed and stained with DAPI. Following a 10 min incubation period at room temperature, stained nuclei were observed under a fluorescence microscope using a blue filter (original magnification 400X). (C) DNA was extracted from cells and analyzed by 1.0% agarose gel electrophoresis in the presence of EtBr. (D) To quantify the degree of apoptotic cells, the cells were collected and stained with PI for flow cytometry analysis. The fraction of apoptotic sub-G1 cells is indicated. Data represent relative mean values ± SD of three independent experiments. The significance was determined by the Student's t-test (*p<0.05, compared with control).
외인적 경로 Death receptor 및 ligand 발현에 미치는 ELM의 영향
Apoptosis 과정은 외인적 경로 및 내인적 경로로 구분되며, 이 과정에서 다양한유전자들의 발현 및 활성 변화와 연계되어 있다. 외인적 경로의 경우는 세포막에 존재하는 DR4, DR5 및 Fas 등과 같은 death receptor에 TRAIL 및 FasL 등과 같은 apoptotic ligand가 결합함으로서 유도된다고 알려져 있다(Kantari and Walczak, 2011; Kaufmann et al., 2011). 이를 위하여 ELM 처리에 따른 apoptosis 유발 과정에서 외인적 경로(extrinsic pathway)를 경유하는 초기단계의 주요 death receptor (DR) 및 ligand 관련 유전자들의 발현 변화를 확인한 결과는 Fig. 2A에 나타난 바와 같다. 결과에서 알 수 있듯이 death receptor 관련 여러 유전자 중 tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL), Fas 및 Fas ligand (FasL)의 발현에는 큰 변화가 관찰되지 않았지만 DR4 및 DR5의 발현은 ELM 처리 농도 의존적으로 증가하였다.
내인적 경로 Bcl-2 family의 발현에 미치는 ELM의 영향
내인적 경로의 경우는 기본적으로 Bcl-2 family 단백질의 발현 변화와 미토콘드리아 막의 기능손상이 동반된다. Bcl-2 family는 apoptosis를 억제하는 anti-apoptotic 유전자들과 apoptosis를 유발하는 pro-apoptotic 유전자들로 구성되어 있으며, 이들은 서로 이합체(dimer)를 형성하며 결합하고 있다(Lee et al., 2018; 2019). 이들 사이의 균형이 깨어지게 되면 미토콘드리아의 기능 이상을 유발하게 되고 미토콘드리아 내막에 존재하고 있는 cytochrome c와 같은 단백질들이 세포질로 방출됨으로서 caspase-9과 최종적으로 caspase-3, -7의 활성화를 통하여 apoptosis가 유발되는 것으로 알려져 있다(Brown et al., 2018; Demine et al., 2019).
내인적 경로(intrinsic pathway)에 있어서 중요한 역할을 하는 것으로 알려진 Bcl-2 family 유전자의 발현에 ELM이 어떠한 영향을 미치는지를 확인한 결과는 Fig. 2B에 나타낸 바와 같이, ELM 처리농도 의존적으로 anti-apoptotic 유전자인 Bcl-2의 발현이 현저하게 감소되었고, pro-apoptotic 유전자인 Bax의 발현은 증가시킴으로써 apoptosis를 유발시킴을 확인할 수 있었다. 또한 활성화된 caspase-8에 의하여 단편화되어 MMP 소실 유발에 중요한 역할을 하는 것으로 알려진 Bid 단백질 발현 변화에도 ELM 처리에 의하여 total Bid의 발현이 감소하는 것으로 나타났으므로 단편화된 형태인 tBid의 증가가 유발되었다는 것을 예상할 수 있었다.
MMP (Δψm) 변화에 미치는 pachymic acid의 영향
Apoptosis 유발에 있어서 미토콘드리아 막의 기능손상이 연관되었을 것으로 추정되어 ELM 처리에 따른 mitochondrial membrane potential (MMP) 값을 dual-emission fluorescent dye인 JC-1 염색으로 통하여 DNA flow cytometer로 분석하였다. Fig. 2C에 나타난 바와 같이 정상배지에서 배양된 U937세포의 MMP 소실 정도는 4.2%로 나타났으나 ELM 처리농도 증가에 따라서 MMP 소실 정도가 18.6%, 36.2%, 56.4%로 현저히 증가되는 것을 확인할 수 있었다.
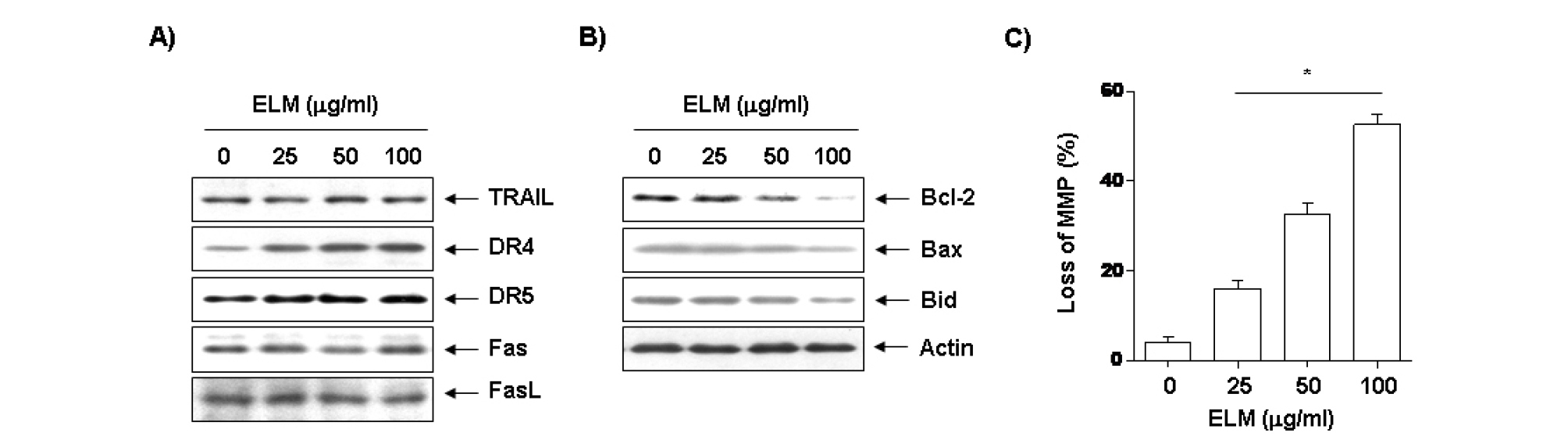
Fig. 2.
Effects of ELM on levels of Death receptor/ligand and Bcl-2 family proteins, and MMP values in U937 cells. (A and B) Following a 24 hr incubation period with various concentrations of ELM, cells were lysed, and cellular proteins were then separated by 10% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. Membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (C) The cells were collected and incubated with 10 μM JC-1 for 20 min at 37℃ in the dark. The cells were then washed once with PBS and mean JC-1 fluorescence intensity was detected using a flow cytometer. Data represent relative mean values ± SD of three independent experiments. The significance was determined by the Student's t-test (*p<0.05, compared with control).
IAP family의 발현에 미치는 SSeo의 영향
Inhibitors of apoptosis protein (IAP) family에 속하는 단백질들은 caspase와 직․간접적으로 결합함으로서 caspase의 활성을 억제하여 apoptosis를 억제하는 것으로 알려져 있다(Cossu et al., 2019; Silke and Vucic, 2014). 이를 위하여 IAP family에 속하는 유전자들의 발현에 ELM이 어떠한 영향을 미치는 지를 확인하였다. Fig. 3A에 나타난 바와 같이, ELM 처리에 의하여 조사된 4가지 IAP family (XIAP, cIAP-1, cIAP-1 및 survivin) 중 XIAP, cIAP-1 및 survivin의 발현이 ELM 처리 농도 의존적으로 감소하였으며, cIAP-2의 발현은 변화가 없음을 확인하였다.
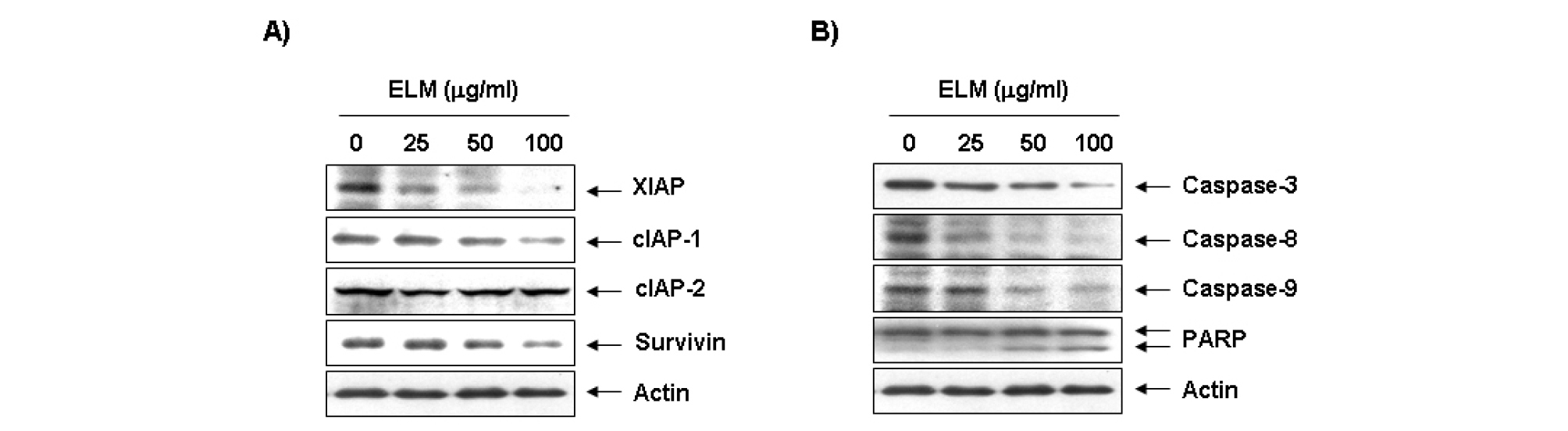
Fig. 3.
Inhibition of IAP family proteins and activation of caspases by ELM in U937 cells. (A and B) After 24 hr incubation with ELM, the cells were collected and lysed. The proteins were then separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
Caspases의 활성에 미치는 ELM의 영향
Caspase는 initiator caspase인 caspase-8 및 -9와 effector caspase인 caspase-3, -6 및 -7 등으로 나누어지는데 먼저 initiator caspase가 활성화 되면 하위단계에 있는 effector caspase를 활성화시킴으로서 세포의 성장 및 생존에 중요한 역할을 하는 여러 종류의 기질 단백질들은 분해함으로서 apoptosis를 유발하는 것으로 알려져 있다(Nagata, 2018; Seo and Rhee, 2018). 특히 caspase-3에 의하여 분해가 유발되는 대표적인 기질 단백질 중의 하나가 PARP이며, caspase-3가 활성화되면 PARP의 단백질이 단편화되어 효소적 기능의 상실로 인하여 정상적인 DNA 수선 과정이 억제되어 apoptosis를 유도되어져 apoptosis 유발의 생화학적 표시자로 널리 사용이 되고 있다. 이를 확인하기 위하여 apoptosis 유발에 중요한 조절유전자로 작용하는 것으로 알려진 caspases 중 extrinsic 및 intrinsic pathway에 직접적으로 관여하는 caspase-8, -9 및 caspase-3의 발현에 미치는 ELM의 영향을 조사하였다. Fig. 3B의 결과에서 알 수 있듯이 ELM 처리에 의하여 조사된 3가지 caspase의 발현이 상대적으로 증가되었음을 알 수 있었다. 또한 caspase-3의 기질 단백질로서 DNA repair와 genomic stability에 관여하는 poly(ADP-ribose)-polymerase (PARP) 단백질의 단편화가 동반되었음을 확인하였다.
적요
본 연구에서는 부처꽃 에탄올 추출물(ELM)에 대한 항암효능을 알아보기 위하여 인체백혈병 U937 세포의 증식에 미치는 영향과 이와 연관된 apoptosis 유발 여부와 함께 그에 따른 분자생물학적 기전에 대해서 조사하였다. 먼저 ELM 처리에 따른 증식억제 정도를 조사한 결과, ELM 처리 농도 의존적으로 생존율 및 증식억제 현상이 나타났으며, 핵의 형태 변화, DNA 단편화 및 apoptosis 유발에 관하여 조사한 결과 역시 ELM 처리 농도 의존적으로 증가됨을 확인할 수 있었다. ELM 처리에 따른 U937 세포에서의 apoptosis 유발에 있어서 미토콘드리아 막의 기능 손상이 관여하는 지를 확인하기 위하여 MMP의 변화 정도를 확인한 결과, ELM 처리 농도 증가에 따라 MMP의 소실이 증가하는 것을 관찰할 수 있었다. 이러한 MMP의 소실에 가 관여하는 지를 확인하기 위하여 사멸수용체(DR4, 5, Fas) 및 사멸수용체에 결합하는 리간드(FasL, TRAIL)의 발현 변화를 확인한 결과, DR4 및 DR5의 발현이 증가하는 것으로 관찰되었다. 또한 내인적 경로에 관여하는 Bcl-2 family 유전자들의 발현변화를 확인한 결과, Bcl-2 발현 감소 및 Bax의 발현 증가의 변화를 보였으며, Bid 단백질의 발현감소가 나타났으므로 상대적으로 tBid의 생성이 증가되었음을 추측할 수 있었다. 한편 apoptosis 유발에 직접적으로 관여하는 것으로 알려진 caspase-3, -8 및 -9의 발현에 미치는 ELM의 영향에 대해서 조사하였다. 결과에서 알 수 있듯이 ELM은 death receptor에 의하여 활성화 되는 것으로 알려진 caspase-8 및 세포질로 방출된 cytochrome c에 의하여 활성화 되는 것으로 알려진 caspase-9의 활성화를 유발하였으며, caspase cascade에 의하여 apoptosis에 직접적으로 관여하는 caspase-3의 발현도 증가시키는 것으로 나타났다. 또한 활성화된 caspase-3에 의하여 분해가 일어나는 기질 단백질인 PARP의 경우 ELM 처리에 의하여 모두 단편화가 유발되는 것으로 나타났다. 이상의 결과를 종합해 보면 인체 백혈병 U937 세포에 ELM을 처리하였을 경우에 유발되는 apoptosis는 외인적 경로인 DR4 및 DR5의 발현 증가를 통한 caspase-8의 활성화와 이로 인한 Bid 단백질의 단편화와 함께 내인적 경로의 미토콘드리아 기능 손실에 의하여 caspase-9 및 -3의 활성화 유발과 기질단백질들의 분해가 중요한 역할을 하는 것으로 생각되며, IAP family의 발현 감소로 인하여 caspase의 활성이 억제되지 못하는 것도 apoptosis 유도에 어느 정도 관여했을 것으로 생각된다. 따라서 ELM 처리에 의하여 유발되는 apoptosis는 외인적 경로 및 내인적 경로를 모두 경유하는 multiple apoptotic pathway에 의하여 조절되며, 이때 caspases가 중요한 역할을 한다는 것을 알 수 있었다.
