

서 언
큰금매화(Trollius macropetalus (Regel) F.Schmidt)는 미나리아재비과(Ranunculaceae)의 금매화 속(Trollius L.)에 속하며, 북반구 온대 및 한대 지방에 걸쳐 분포하고 있다. 전 세계적으로 35종이 Trollius 속에 속해있으며(Luferov et al., 2018), 주로 산림 스텝, 아고산 및 고산지 등에 서식하고 있다(Serebryanyi, 2019). 한반도에 자생하는 Trollius 속은 큰금매화, 애기금매화(T. riederianus Fisch. and C.A.Mey.), 금매화(T. ledebourii Rchb.)로 총 3종이 보고되었다(Park and Lee, 2018). Trollius 속은 전통 민간 약재로 널리 사용되어 왔다. 북금매화(T. chinensis Bunge.)는 항박테리아, 항바이러스, 항산화 및 항종양 효과가 있는 것으로 알려져 있어(Jiang et al., 2020; Li et al., 2002; Lu et al., 2015; Yang et al., 2013) 근연종인 큰금매화의 의약적 가치 또한 주목된다.
큰금매화는 고산 초지에 주로 서식하는 여러해살이풀이다. 초장은 30~60 ㎝이며 7~8월에 개화한다. 타원형의 꽃받침이 5~7개이고, 선상 피침형의 꽃잎이 수술보다 긴 것이 특징이다(Lee, 2003). 지름 4 ㎝의 등황색 꽃이 줄기와 가지 끝에 1개씩 달리며, 꽃의 관상적 가치가 뛰어나 조경용 소재로 주목받고 있다(Song et al., 2015). 큰금매화의 분포지는 중국 만주 지방과 극동 러시아, 한반도 북부인 북한 양강도, 함경남도, 자강도로 남한에서는 자생지가 보고된 적이 없다. 또한 큰금매화는 북한에서도 부전고원과 같은 고산지에 서식하고 있는 고산성 북방계 식물로서 기후 변화에 특히 취약한 종으로 알려져 있다(Gantsetseg et al., 2020). 현재 북한은 고질적인 식량난과 자연재해로 인해 산림황폐화 및 생물종다양성 감소가 심각하며(DPR of Korea, 2007; Park and Yoo, 2009), 남한에는 큰금매화가 분포하지 않아 유전자원의 확보가 어려운 상황을 고려하였을 때 큰금매화 유전자원의 현지 외(ex-situ) 보전이 절실한 상황이다. 식물의 종자는 보관이 용이하여 연구 및 재도입, 복원 소재로의 활용도가 높아 현지 외(ex-situ) 보전의 중요 방법으로 여겨지고 있다. 하지만 이러한 종자의 활용을 위해서는 종자의 휴면타파 및 발아 조건에 관한 연구가 필수적으로 요구된다(Hay and Probert, 2013; Kim et al., 2023; O'Donnell and Sharrock, 2017; Suh et al., 2022).
종자 휴면은 식물 개체가 생존에 불리한 환경에서 종자 발아를 억제하여 생존율을 높이기 위한 전략으로, 기작에 따라 생리적 휴면(PD, Physiological dormancy), 형태학적 휴면(MD, Morphological dormancy), 물리적 휴면(PY, Physical dormancy), 형태·생리학적 휴면(MPD, Morpho-physiological dormancy), 조합휴면(PY+PD, Combinational dormancy)의 5가지 유형으로 나뉜다(Baskin and Baskin, 2014; Nonogaki, 2019). 큰금매화가 속한 미나리아재비과 식물의 종자 휴면 유형은 주로 탈리 시 직선형의 미숙배를 지닌 MD 혹은 MPD로 보고되었고(Baskin and Baskin, 2014), 금매화 종자 또한 직선형과 미발달형의 배를 지닌 것으로 보고되었다(Hepher and Roberts, 1985). 큰금매화의 종자 휴면 유형에 대한 직접적인 연구는 이루어지지 않았지만, 금매화 종자의 경우 GA4+7를 처리했을 때 발아율이 3배 이상 증가하였다고 보고되었다(Hepher and Roberts, 1985). 20℃ 이상의 실온에서 배양시 금매화는 22일의 휴면 기간과 92%의 발아율, 북금매화는 70~110일의 휴면 기간과 93.5%의 발아율을 나타낸 반면, T. apertus와 T. europaeus, T. asiatirux × T. kytmanovii는 약 3~10%의 낮은 발아율을 보인다고 하였다(Gusar et al., 2021).
현재 북한지역 내 분포하는 식물 중 증식 및 번식에 관한 연구는 일부 분류군에서만 이루어졌고, 큰금매화 또한 종자의 휴면 유형 및 발아 조건에 관한 연구는 이루어지지 않았다. 이에 본 연구에서는 큰금매화 종자의 휴면 유형 구명을 위해 변온 조건, 저온층적처리 및 지베렐린(GA3) 처리가 발아에 미치는 영향을 조사하였다.
재료 및 방법
공시 재료
실험에 공시한 큰금매화(T. macropetalus (Regel) F.Schmidt) 종자는 국립DMZ자생식물원에서 저장 중이던 종자를 사용하였다. 큰금매화 종자는 2021년 국립DMZ자생식물원에서 채종되어 통풍이 잘 되는 곳에서 상온 건조되었다. 이후 온도 4℃, 상대습도 40%로 1년간 저장된 것을 정선하여 형태·생리적 특성조사와 발아 검정에 사용하였다.
종자 형태·생리적 특성조사
큰금매화 종자의 외부 형태를 관찰하기 위하여 디지털현미경(DVM6, Leica Microsystems GmbH, Wetzlar, Germany)을 이용하여 종자의 크기와 형태를 관찰하였다. 종자의 활력은 국제종자검정협회(ISTA, International Seed Testing Association)의 Tetrazolium 검정 방법에 준하여 조사하였다(ISTA, 2014). 종자 상단에 면도날을 이용하여 얇은 칼집을 내고 증류수에 침지한 다음(20℃, 18시간), 2, 3, 5-Triphenyl tetrazolium chloride 1% 용액에 침지하였다(30℃, 18시간). 침지 후 종단면을 잘라 해부현미경(SMZ1500, Nikon, Tokyo, Japan)으로 종자 내부 배의 염색 정도와 상태를 관찰하였고, X-ray 검사기(EMT-F70, Softex, Japan)를 이용하여 종자의 활력을 분석하였다.
온도처리 및 발아조사
큰금매화 종자를 500 ㎎·L-1의 소독제(Benomyl, FarmHannong, Seoul, Korea)에 1시간 침지하여 소독한 뒤 증류수로 4회 이상 수세한 후 치상하였다. 처리구별 90×15 ㎜ Petri dish를 사용하였고, 0.8% Agar 배지 위에 종자를 30립씩 치상하였다. 치상된 큰금매화 종자는 처리구별 3 반복으로 완전임의배치하였다.
온도에 따른 큰금매화 종자의 발아 반응을 알아보기 위해 치상된 종자를 15/6℃, 25/15℃ 변온(12/12 시간)으로 조절된 생장상(WCC-1000, Daihan Scientific Co., Wonju, Korea)에서 배양하였다. 생장상 내 광도 조건은 10±2 μmol·m-2·s-1이었고, 하루 16시간 명조건을 유지하였다.
유근이 종피를 뚫고 1 ㎜ 이상 돌출된 종자를 발아한 것으로 간주하였다. 치상 후 24시간 간격으로 발아 개체수를 조사하였고, 총 30일 동안 조사하였다. 조사 기간 중 발아된 종자들은 제거하였고, 부패한 종자들은 발아율 계산에서 제외하였다. 조사 결과를 바탕으로 큰금매화 종자의 최종발아율(FGR, Final germination rate), 평균발아일수(MGT, Mean germination time), 발아균일도(GU, Germination uniformity), 발아세(GS, Germination speed)를 아래의 식을 이용하여 산출하였다(Ellis, 1981).
· FGR = (N / S) × 100
· MGT = Σ (Tx · Nx) / N
· GU = Σ [(MGT - Tx)2 · Nx] / (N - 1)
· GS = Σ (Nx / Tx)
· (N: 총 발아수, S: 총공시 종자수, Nx: 조사 당일의 발아수, Tx: 치상 후 조사일수)
저온층적처리
저온에 의한 휴면타파 효과를 알아보기 위하여 큰금매화 종자를 대상으로 저온층적처리를 실시하였다. 온도처리와 동일한 방법으로 종자를 소독 및 치상하였고, Petri dish 내 수분 손실을 막기 위해 Parafilm으로 밀봉한 뒤 Aluminum foil을 두 겹 감싸 암처리하였다. 4℃의 항온 생장상(WIM-RL4, Daihan Scientific Co., Wonju, Korea)에서 6주간 처리한 후 15/6℃, 25/15℃의 변온으로 조절된 생장상(WCC-1000, Daihan Scientific Co., Wonju, Korea)에서 배양하였다. 생장상 내 광도 조건은 온도처리구와 동일하였다.
지베렐린 처리
GA3 처리에 의한 휴면타파 효과를 알아보기 위하여 큰금매화 종자를 대상으로 GA3을 처리하였다. 온도 및 저온층적처리와 동일한 방법으로 종자를 소독하였고, 소독한 종자를 500과 1,000 ㎎·L-1의 GA3 용액에 24시간 동안 암조건에서 침지하였다. 침지 처리된 종자를 증류수로 5회 수세한 뒤 온도처리와 동일한 방법으로 Agar 배지에 치상하였고, 15/6℃, 25/15℃의 변온으로 조절된 생장상(WCC-1000, Daihan Scientific Co., Wonju, Korea)에서 배양하였다. 생장상 내 광도 조건은 온도처리구와 동일하였다.
통계처리
변온 조건 및 GA3 처리 실험에 따른 큰금매화 종자의 발아 특성 차이를 SPSS 프로그램(version 12.0, IBM Inc., USA)으로 분산분석(ANOVA)하였고, Scheffe’s multiple range test (p < 0.05)로 처리별 평균 차이의 통계적 유의성을 검정하였다.
결 과
종자 형태·생리적 특성
큰금매화 종자는 원형∼계란형(Circular ~ Obovate)에 표면에 작은 돌기가 겹겹이 있는 형태(Colliculate)였으며, 종피는 진갈색이다(Fig. 1a and 1b). 큰금매화 종자의 크기는 길이 1.37±0.74 ㎜, 너비 0.81±0.53 ㎜이고, 천립중은 11.28±0.72 g이었다. Tetrazolium 검정 결과에서 모든 종자의 배가 전체적으로 붉게 염색되어 종자의 활력은 100%로 나타났고(Fig. 1c), X-ray 검정 결과에서 큰금매화 종자는 91%의 충실도를 보였다(Fig. 1d). 발아가 시작되면 유근이 주공을 통해 나타났고, 떡잎이 완전히 출현하기까지 하루에서 이틀정도 소요되었다(Fig. 2).

Fig. 1.
(a) Seed morphology of Trollius macropetalus (Regel) F.Schmidt, (b) seed morphology observed by electron microscope (SEM), (c) undeveloped embryos and endosperm stained with tetrazolium are shown in the initial seed at seed coat split. EM, embryo; ES, endosperm; SC, seed coat. (d) Seed fullness of T. macropetalus via X-ray inspection.
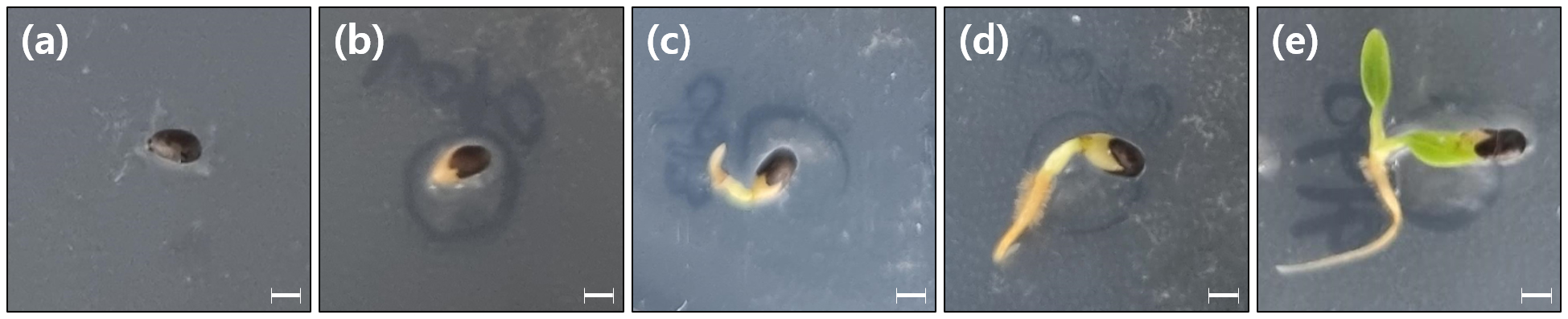
처리별 종자 발아 특성
무처리한 큰금매화 종자의 발아 특성을 살펴보면, 15/6℃ 변온 조건에서의 최종발아율이 12.2±3.1%로 나타났다(Table 1). 또한 평균발아일수는 22.9±0.7일이고, 발아균일도는 2.5±1.5, 발아세는 0.2±0.1로 나타났다. 한편 25/15℃ 변온 조건에서는 큰금매화 종자의 발아를 확인할 수 없었다.
6주간 4℃로 저온층적처리된 큰금매화 종자는 15/6℃ 변온 조건에서 1.1±1.6%의 최종발아율을 나타냈고, 평균발아일수는 4.7±3.1일, 발아균일도와 발아세는 0.0±0.0로 나타났다(Table 1). 25/15℃ 변온 조건에서는 최종발아율이 2.2±3.1%로 나타났고, 평균발아일수가 3.8±3.6일, 발아균일도는 4.2±3.2, 그리고 발아세는 0.1±0.1로 나타났다. 저온층적처리된 큰금매화 종자의 최종발아율은 두 변온 조건 간에 유의미한 차이를 보이지 않았고, 15/6℃ 변온 조건에서의 최종발아율은 무처리구에 비해 현저히 저하하였다. 평균발아일수와 발아균일도 또한 유의미한 차이가 없었고, 발아균일도는 무처리구와 유의미한 차이가 없었던 반면 평균발아일수는 무처리구에 비해 4배 이상 단축되었다. 발아세는 15/6℃보다 25/15℃ 변온 조건에서 더 높았으나 두 조건 모두 무처리구에 비해 유의미하게 낮았다.
GA3가 처리된 큰금매화 종자의 발아 특성은 다음과 같이 나타났다(Table 1). 15/6℃ 변온 조건에서의 결과를 살펴보면 GA3 500 ㎎·L-1 처리구의 최종발아율은 36.7±4.7%로 나타났다. 그리고 평균발아일수는 16.8±1.8일, 발아균일도는 22.6±7.7, 발아세는 0.7±0.0로 나타났다. GA3 1,000 ㎎·L-1 처리구의 최종발아율은 40.0±14.1%로 나타났고, 평균발아일수 16.9±0.7일, 발아균일도 14.5±5.4, 발아세는 0.8±0.4였다. 25/15℃ 변온 조건에서는 GA3 500 ㎎·L-1 처리구의 최종발아율이 27.8±4.2%, 평균발아일수가 14.6±0.9일, 발아균일도는 20.1±4.5, 발아세는 0.6±0.1로 나타났다. GA3 1,000 ㎎·L-1 처리구의 최종발아율은 33.3±9.8%로 나타났고, 평균발아일수 15.6±2.0일, 발아균일도 15.8±5.5, 발아세는 0.7±0.3이었다. 15/6℃ 변온 조건에서 GA3가 처리된 큰금매화 종자의 최종발아율은 500 ㎎·L-1처리구와 1,000 ㎎·L-1처리구 간에 유의미한 차이가 없었고, 평균발아일수와 발아균일도 또한 마찬가지였다. 한편 발아세는 500 ㎎·L-1처리구에 비해 1,000 ㎎·L-1처리구가 더 높았다. 25/15℃ 변온 조건에서의 최종발아율은 500 ㎎·L-1처리구에 비해 1,000 ㎎·L-1처리구에서 현저히 증가하였으나 평균발아일수와 발아균일도 간의 유의미한 차이가 없었다. 발아세는 15/6℃ 변온 조건과 마찬가지로 500 ㎎·L-1처리구에 비해 1,000 ㎎·L-1처리구에서 더 높았다. 15/6℃ 변온 조건에서 GA3 500 ㎎·L-1및 1,000 ㎎·L-1 처리구의 최종발아율은 무처리구와 저온층적처리에 비해 현저히 증가하였다. 반면 25/15℃ 변온 조건에서는 1,000 ㎎·L-1 처리구의 최종발아율만이 무처리구와 저온층적처리에 비해 현저히 증가하였고, 500 ㎎·L-1 처리구의 최종발아율은 15/6℃ 변온 조건의 무처리구와 유의미한 차이가 없었다. 평균발아일수는 GA3 처리구 간에 유의미한 차이가 없었으나 무처리구에 비해 단축되고 저온층적처리에 비해 지연되는 결과를 보였다. 모든 GA3 처리구의 발아균일도는 무처리구, 그리고 저온층적처리와 유의미한 차이를 보이지 않았다. 25/15℃ 변온 조건에서 GA3 500 ㎎·L-1처리구의 발아세는 15/6℃ 변온 조건의 무처리구와 유의미한 차이가 없었으나 이를 제외한 GA3 처리구들의 발아세는 무처리구와 저온층적처리에 비해 현저히 증가하였다.
Table 1.
Final germination rate (FGR), mean germination time (MGT), germination uniformity (GU), and germination speed (GS) of Trollius macropetalus (Regel) F.Schmidt seeds under controlled temperature conditions (15/6℃ and 25/15℃) and pretreatments.
| Treatment | FGR (%) | MGT (day) | GU | GS |
| 15/6℃ NTz | 12.2 ± 3.1abx | 22.9 ± 0.7a | 2.5 ± 1.5a | 0.2 ± 0.1ad |
| 15/6℃ CSy | 1.1 ± 1.6b | 4.7 ± 3.1bc | 0.0 ± 0.0a | 0.0 ± 0.0cd |
| 15/6℃ GA3 500 ㎎·L-1 | 36.7 ± 4.7a | 16.8 ± 1.8ab | 22.6 ± 7.7a | 0.7 ± 0.0ac |
| 15/6℃ GA3 1000 ㎎·L-1 | 40.0 ± 14.1a | 16.9 ± 0.7ab | 14.5 ± 5.4a | 0.8 ± 0.4a |
| 25/15℃ NT | 0.0 ± 0.0b | 0.0 ± 0.0c | 0.0 ± 0.0a | 0.0 ± 0.0d |
| 25/15℃ CS | 2.2 ± 3.1b | 3.8 ± 3.6bc | 4.2 ± 3.2a | 0.1 ± 0.1bd |
| 25/15℃ GA3 500 ㎎·L-1 | 27.8 ± 4.2ab | 14.6 ± 0.9ab | 20.1 ± 4.5a | 0.6 ± 0.1ad |
| 25/15℃ GA3 1000 ㎎·L-1 | 33.3 ± 9.8a | 15.6 ± 2.0ab | 15.8 ± 5.5a | 0.7 ± 0.3ab |
처리별 최초 발아일과 최종 발아일을 살펴보면 다음과 같다(Fig. 3). 15/6℃ 변온 조건에서 무처리된 큰금매화 종자는 21.7±0.9일에 처음 발아하여 24.3±0.9일까지 발아가 지속되었다. 저온층적처리된 큰금매화 종자는 14.0±0.0일에 발아하여 이후부터 발아가 관찰되지 않았다. GA3 500 ㎎·L-1 처리된 조건에서는 10.7±0.5일부터 24.7±1.9일까지 발아가 지속되었고, 1,000 ㎎·L-1 처리된 조건에서는 14.0±2.8일부터 22.7±2.9일까지 발아가 지속되었다. 25/15℃ 변온 조건에서는 저온층적처리된 큰금매화 종자가 9.0±0.0일에 처음 발아하여 14.0±0.0일까지 발아가 지속되었다. GA3 500 ㎎·L-1 처리된 조건에서는 10.3±0.5일부터 22.3±2.9일까지 발아가 지속되었고, 1,000 ㎎·L-1 처리된 조건에서는 11.3±1.9일부터 20.0±4.1일까지 발아가 지속되었다.

Fig. 3.
Cumulative germination rate from Trollius macropetalus (Regel) F.Schmidt pretreated with GA3 and incubated under temperature conditions of (a) 15/6℃ and (b) 25/15℃. There was no significant difference between the cold stratification treatment (CS) and the non-treatment (NT) at 25/15℃, but a significant difference was found at 15/6℃. Error bar means the standard deviation.
고 찰
종자는 내부에 위치한 배의 형태에 따라서 기저형(Basal), 지엽형(Peripheral) 그리고 증축형(Axile)의 세 가지로 나뉜다(Martin, 1946). 증축형은 배의 크기가 작은 것부터 큰 것까지 다양하며, 배가 종자의 중앙에 길게 위치한 선형(Linear-type)이 가장 대표적이다(Song et al., 2019). 본 연구 결과, 큰금매화 종자의 배 형태는 작은 선형(Axial-miniature)으로 판단된다(Fig. 1c). 증축형(Axile)의 종자는 길이에 따라 구분할 수 있고, 길이 0.3~2.0 ㎜는 Dwarf-type, 0.2 ㎜ 이하는 Micro-type으로 분류된다(Martin, 1946). 이에 따라 큰금매화 종자를 Micro-type으로 판단한다.
미성숙 배는 적절한 온도에서 성숙하여 유근을 형성하고, 그 후 일정 기간의 2차 배축 휴면을 통해 떡잎을 형성한다. 이처럼 미숙배를 가진 종자는 발아 전에 배가 일정 크기 이상으로 자라야 한다. 배가 적절한 조건에서 30일 이내에 신장과 발아가 이루어지면 형태적휴면(MD), 발아하는데 시간이 30일 이상 소요되거나 저온층적처리 또는 기타 생리활성물질을 처리해야 발아하는 경우를 형태·생리적휴면(MPD)으로 분류한다(Baskin and Baskin, 2004). 큰금매화의 종자는 발아하는데 30일 이상 소요되지 않아 형태적휴면으로 보이나(Fig. 3) 무처리구의 발아율이 15/6℃에서 12.2±3.1%에 그쳤고, 25/15℃에서는 전혀 발아하지 않았다(Table 1). 한편 25/15℃ 무처리구에서는 전혀 발아하지 않았음에도 GA3 처리로 발아한 점으로 미루어 볼 때 큰금매화는 MD보다 MPD가 강하게 나타나는 것으로 추정된다. 이는 미나리아재비과 식물의 종자 휴면이 2가지 유형으로 보고된 것과 유사한 결과였다(Baskin and Baskin, 2014). 종자의 휴면은 일반적으로 종 특이적인 기작이지만 같은 속 내에서도 생태적 특성과 지리적 분포 때문에 종류와 정도가 다르게 나타나기도 한다(Park et al., 2019). 미나리아재비과 Trollius 속 식물들의 종자 휴면과 발아율은 종간 차이가 있으며, 주로 MD 혹은 MPD를 지닌다고 하였고(Gusar et al., 2021), 큰금매화의 근연종인 금매화는 MPD를 가진다(Hepher and Roberts, 1985).
금매화는 늦은 봄에 미성숙 배로 탈립된 다음 그해 여름과 겨울을 나면서 휴면이 타파된다(Lee et al., 2003). 큰금매화는 7~8월에 개화하여 가을에 결실된다는 점에서 금매화와 차이가 있지만 모수에서 탈리된 종자가 노지에서 월동해야 한다는 점은 동일하다. 미성숙 배의 후숙이나 휴면타파에는 온도가 주요 요인으로 알려져 있으며(Lee et al., 2003), MPD는 배 발달의 온도 범위가 15~20℃인 Simple-type과 0~10℃인 Complex-type으로 구분할 수 있다(Baskin and Baskin, 1998, 2004; Geneve, 2003; Song et al., 2019). 비록 온도 범위에 따라 종자 내부의 미숙배가 얼마나 신장하였는지 본 연구 결과에서는 제시하지 못했지만 무처리구 기준 15/6℃ 변온 조건에서만 발아가 이루어졌기 때문에 Complex-type으로 생각된다.
형태·생리적휴면은 형태적휴면과 생리적휴면이 복합적으로 작용하고, 생리적휴면(PD)의 깊이에 따라서 Deep-type, Intermediate-type, Nondeep-type으로 구분할 수 있다(Baskin and Baskin 1998, 2004; Geneve, 2003). Deep-type PD는 GA 처리로 휴면이 타파되지 않으며, 휴면을 타파하는데 3~4개월의 저온층적처리 기간이 필요하다. Intermediate-type PD 또한 휴면을 타파하는데 2~3개월의 저온층적처리 기간이 필요하고, 일부 종들에서만 GA 처리가 휴면타파에 효과적이다. Nondeep-type PD는 짧게는 수일에서 길게는 2개월 정도의 저온층적처리로 휴면이 타파되며, GA 처리로 휴면이 쉽게 타파되고 후숙 효과 또한 있다고 알려져 있다(Song et al., 2019). GA3는 종자의 휴면을 타파하고 발아를 촉진하는 식물호르몬으로, 휴면을 유발하는 ABA와 상반되는 역할을 한다(Jang et al., 2016; Kim and Lee, 2013). GA/ABA 비율과 민감도는 종자의 휴면과 발아의 균형을 조절하는 요소로 알려져 있으며(Cho et al., 2018; Finch-Savage and Leubner-Metzger, 2006), GA 함량이 증가하여 ABA에 대한 민감도가 감소하면 휴면이 타파되어 발아가 촉진되는 현상이 나타난다(Song et al., 2019). 본 연구 결과에서 큰금매화의 종자는 저온층적처리로 휴면이 타파되지 않았으나 GA3 처리구의 발아율이 무처리구에 비해 현저히 증가하였기 때문에 Nondeep-type으로 판단된다.
이상의 결과로부터 큰금매화는 15/6℃ 변온 조건과 GA3 500~1,000 ㎎·L-1 처리시 종자 발아가 가장 우수한 것으로 확인되었다. 큰금매화는 두 변온 조건에서 20% 미만의 발아율을 나타냈으나 GA3 처리 시 휴면이 타파되어 nondeep complex-type MPD로 판단된다. 본 연구에서는 배의 성숙에 관여하는 기타 생리활성물질과 해부학적 관찰에 관한 연구가 미비하였다. 향후 연구에서 GA3 뿐만 아니라 생장조절제와 무기염류 처리가 큰금매화의 발아에 미치는 영향을 조사하고, 종자 길이에 대한 배의 크기를 상대적으로 표현할 수 있는 E:S ratio를 면밀히 조사한다면 발아 기작에 대한 정보를 보완할 수 있을 것이다. 큰금매화 종자가 고온 환경에서 발아하지 않는 것은 큰금매화의 자생지가 고산지에만 분포한다는 사실과 연관이 있어 보인다. 이를 명확히 구명하기 위해 고온습윤처리(Warm stratification)나 항온 조건이 발아에 미치는 영향을 후속 연구로 진행하여야 할 것이다. 이 결과는 큰금매화의 종자 발아에 관한 첫 번째 연구이고, 향후 형태·생리학적 발아 메커니즘을 구명하는데 필요한 자료가 될 것으로 기대한다. 또한 이는 북방계 식물인 큰금매화를 현지내·외 보전하는데 중요한 자료가 될 것이다.
적 요
본 연구는 북한 식물자원 큰금매화의 종자 번식을 위해 발아에 대한 기초자료를 제시하고자 수행되었다. 큰금매화 종자의 형태적 특성과 활력을 검정하였고, 변온 조건(15/6℃ & 25/15℃), 저온층적처리 및 GA3 처리에 따른 발아 특성을 조사하였다. 연구 결과, 저온층적처리는 큰금매화 종자의 휴면타파에 효과가 없었다. GA3 처리는 큰금매화 종자의 발아율을 현저히 증가시켰고, 평균발아일수와 발아세 또한 유의미하게 향상시켰다. 한편, 15/6℃ 조건에 비해 25/15℃ 조건에서는 큰금매화 종자에 대한 GA3 처리의 효과가 감소하였다. 큰금매화 종자는 Nondeep-type의 형태·생리적휴면(MPD)을 가지는 것으로 판단되었고, 본 연구에서 가장 효율적인 발아 조건은 15/6℃ 변온 조건과 GA3 500 ㎎·L-1 처리로 확인되었다. 이러한 본 연구의 결과는 고산성 북방계 식물인 큰금매화의 대량증식에 유용한 자료가 될 것이다.
