
-
Research Article
- Immature Persimmon (Diospyros kaki Thunb.) Ethanol Extract Inhibits Adipogenesis through AMP-Activated Kinase in 3T3-L1 Preadipocytes
- Seon-A Yoon, Hyejin Hyeon, Ho Bong Hyun, Sung Chun Kim, Boram Go, Soyeon Oh, Ji-Gweon Park, Young-min Ham
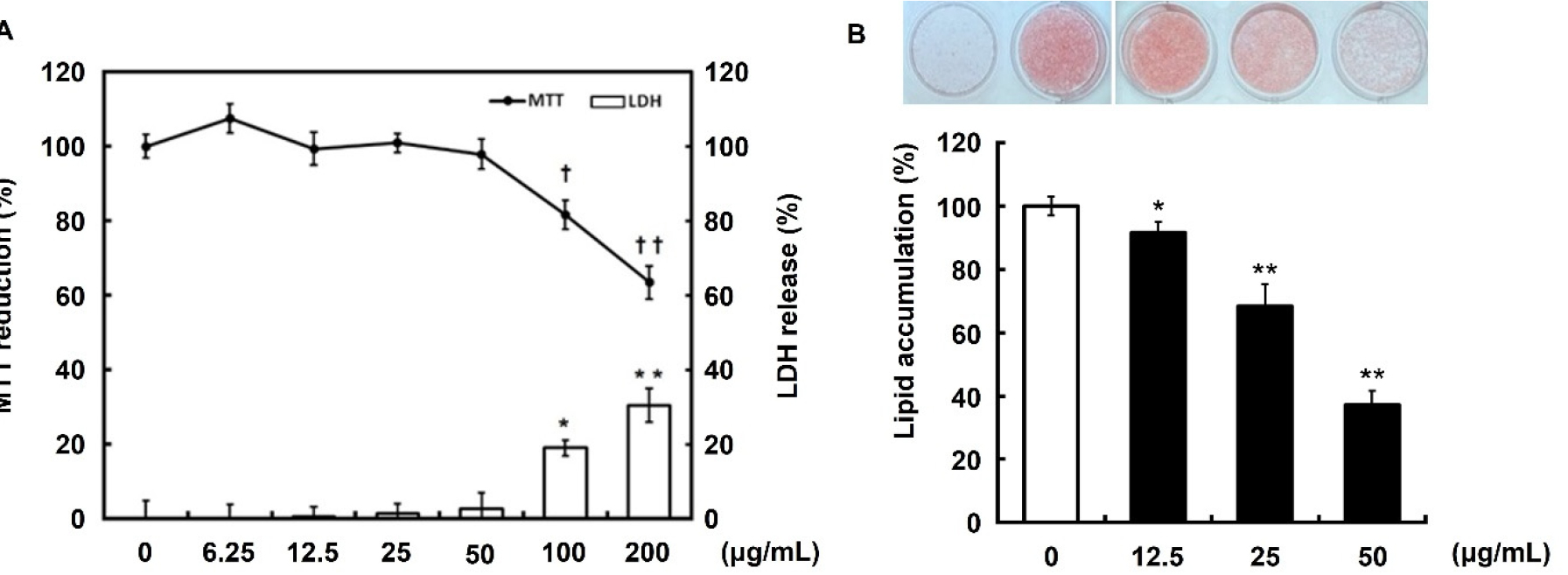
- We previously reported the anti-obesity effects of immature persimmon ethanol extract by analyzing changes in body weight, visceral fat weight, blood biochemicals, …
- We previously reported the anti-obesity effects of immature persimmon ethanol extract by analyzing changes in body weight, visceral fat weight, blood biochemicals, and fat production-related gene expression levels in a high-fat diet-induced obese mouse model. Therefore, this study investigated whether the 50% ethanol extract of Diospyros kaki Thunb. regulated lipid metabolism by identifying its mechanism in 3T3-L1 cells. As a result, it did not influence cell viability or toxicity, and a significant decrease in lipid droplet accumulation was observed in a concentration-dependent manner during adipogenesis (adipogenic differentiation). Consistent with the animal studies, Diospyros kaki Thunb. extract (in a dose-dependent manner) suppressed peroxisome proliferator-activated receptor γ and CCAAT/enhancer-binding proteins, which induce adipocyte differentiation in 3T3-L1 preadipocytes. In addition, it increased the levels of phosphorylated AMP-activated protein kinase (AMPK) and acetyl-CoA carboxylase in a dose-dependent manner in 3T3-L1 cells. Finally, we confirmed that sterol regulatory element-binding protein-1c is regulated by AMPK phosphorylation. These results indicated that immature persimmons (Diospyros kaki Thunb.) may serve as a natural agent for the prevention or treatment of obesity via the AMPK pathway. - COLLAPSE
-
Research Article
- Variations in Vitamin C, Fatty Acids, Total Metabolites, and Antioxidant Capacity Levels of Yardlong Bean Accessions Grown in Korea
- Kebede Taye Desta, Myoung-Jae Shin, Hyemyeong Yoon, Jungyoon Yi, Sukyeung Lee, Yu-Mi Choi
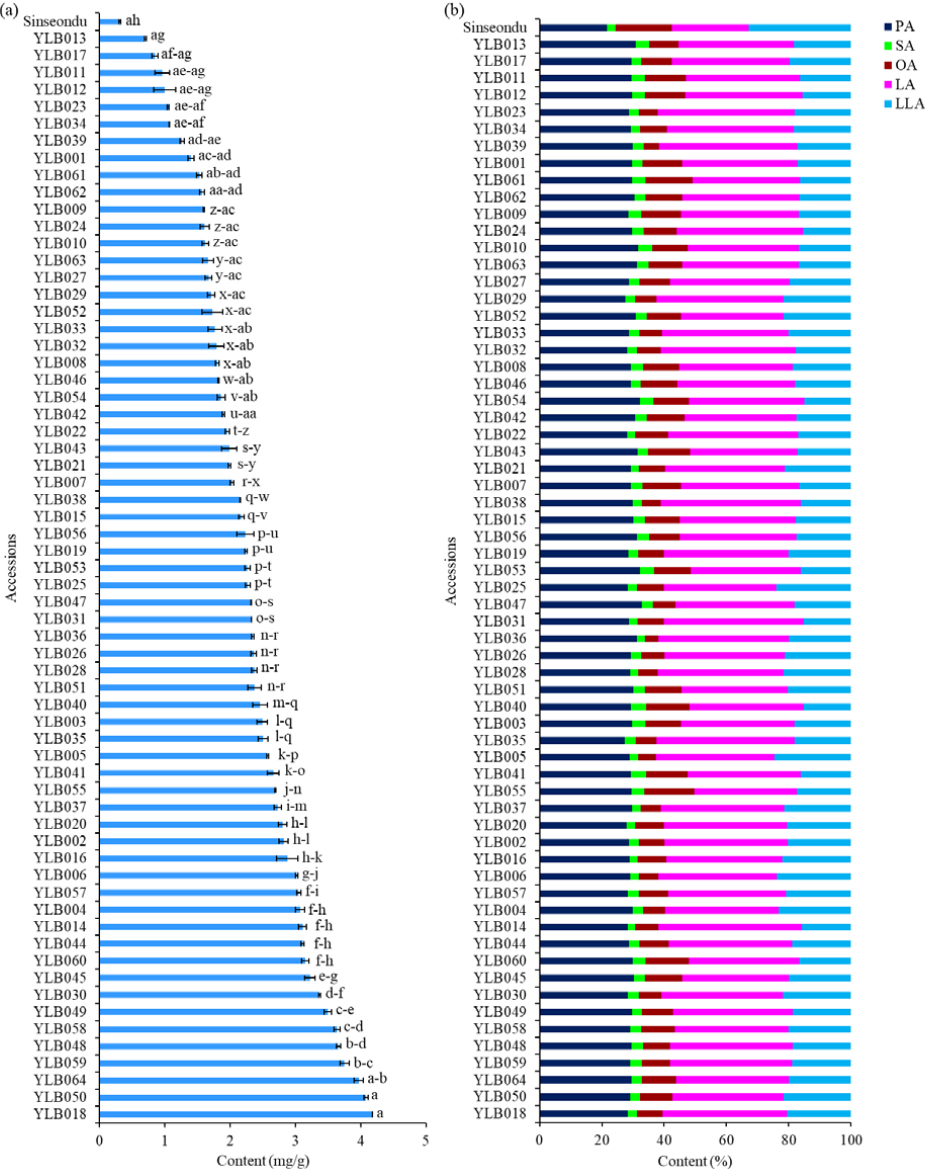
- This study examined the variations of key biochemical components across 64 yardlong bean accessions grown in Korea and compared their performance with …
- This study examined the variations of key biochemical components across 64 yardlong bean accessions grown in Korea and compared their performance with Sinseondu, a local common bean cultivar. All parameters showed significant variations (p < 0.05), indicating a wide genetic diversity among the accessions. Vitamin C content ranged from 0.31 to 4.18 ㎎/g and exhibited a coefficient of variation (CV) of 37.23%. Likewise, the contents of palmitic acid, stearic acid, oleic acid, linoleic acid, and linolenic acid were in the ranges of 21.69–32.84, 2.30–4.87, 4.23–18.04, 24.66–46.02, and 15.03–32.83%, respectively. The CV for these fatty acids ranged from 5.02% in palmitic acid to 28.01% in oleic acid, signifying lipid quality differences. Total phenol, total tannin, and total saponin contents showed more than three-fold variation, while antioxidant activities, including DPPH• scavenging activity, ABTS•+ scavenging activity, and reducing power, showed over six-fold variation. The control cultivar had the lowest levels of vitamin C, total phenol, palmitic acid, and linoleic acid, but the highest linolenic acid. Overall, Sinseondu exhibited a better lipid quality, but lower antioxidant properties than the yardlong bean accessions. Cluster analysis divided the studied accessions into seven groups, and showed significant variation in all parameters (p < 0.05). Principal component and correlation analyses also showed noteworthy associations between the analyzed parameters. This study demonstrated the biochemical diversity of yardlong beans recently cultivated in Korea. Accordingly, sixteen accessions with desirable qualities were identified and hence, they could be used in food industries and breeding programs. - COLLAPSE
-
Research Article
- Shading Treatment-Induced Changes with Growth and Physiological Characteristics in Delphinium maackianum Regel
- Seungju Jo, Dong-Hak Kim, Jung-Won Yoon, Eun Ju Cheong
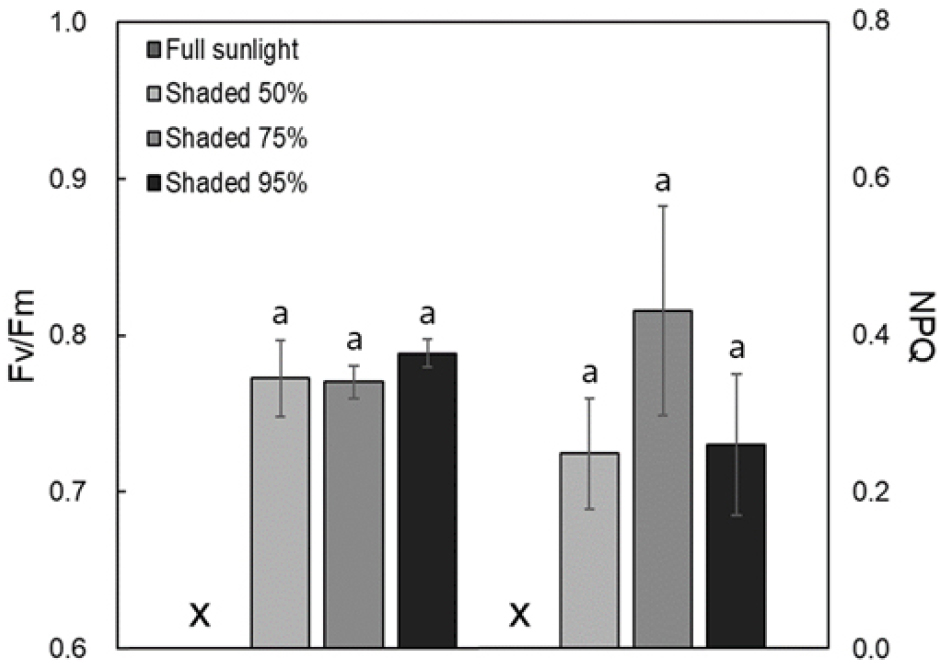
- Delphinium maackianum was grown under different shade treatments to examines the impact of different shading levels on the growth, physiological characteristics. The …
- Delphinium maackianum was grown under different shade treatments to examines the impact of different shading levels on the growth, physiological characteristics. The leaf mass per area (LMA) was lowest in the 95% shading treatment, indicating thinner and broader leaves, however significant differences were not observed across shading levels. Chlorophyll content (a, b, and total) increased with higher shading. Fv/Fm and NPQ values showed no significant differences among treatment groups except for the full light treatment, suggesting maintained photosystem II functionality and photoprotective mechanisms. The dark respiration rate (Rd) and light compensation point (LCP) decreased with increased shading, reflecting shade-tolerant plant responses. The 95% shading treatment exhibited the highest photosynthetic rates, light saturation points (LSP), and apparent quantum yields (AQY), indicating enhanced photosynthetic efficiency under low light conditions. Correlation analysis revealed negative relationships between relative light intensity and several physiological traits, including chlorophyll content, LCP, LSP, maximum photosynthetic rate, and AQY. This indicates that D. maackianum optimizes photosynthesis under low light conditions by increasing light absorption and utilization efficiency. The study underscores the species’ ability to adjust its physiological processes and structural characteristics to thrive in varying shading environments, highlighting its adaptive strategies for survival and growth in shaded habitats. - COLLAPSE
-
Research Article

- A New Distribution Record of Arthraxon prionodes (Poaceae) in Korea
- Jung-Hyun Kim, Yang-Hoon Cho, Seok-Soon Kim, Sunhee Sim, Min-Ha Kim
- A new distribution of Arthraxon prionodes (Steud.) Dandy is discovered in Korea. This species was collected from rocky mountain slopes on Daeyang-ri, …
- A new distribution of Arthraxon prionodes (Steud.) Dandy is discovered in Korea. This species was collected from rocky mountain slopes on Daeyang-ri, Nami-myeon, Geumsan-gun, Chungcheongnam-do. Arthraxon prionodes is well distinguished from other species of the genus by its perennial habit, lower glume of sessile spikelet laterally 2-keeled with inflexed margins, and 3 stamens. Here, we provide detailed morphological description, taxonomic key to Korean Arthraxon species, voucher specimens and photographs of its habit. - COLLAPSE

Journal Informaiton
 Korean Journal of Plant Resources
Korean Journal of Plant Resources
Korean Journal of Plant Resources







한국자원식물학회
The Plant Resources Society of Korea 6 Jechon-si, Chungbuk 27116, Korea
Tel: +82-31-202-4116 / Fax: +82-0502-202-4567 / E-mail: prsk1001@hanmail.net Copyright© The Plant Resources Society of Korea. Powered by APUB
Tel: +82-31-202-4116 / Fax: +82-0502-202-4567 / E-mail: prsk1001@hanmail.net Copyright© The Plant Resources Society of Korea. Powered by APUB