

서언
유채(Brassica napus L.)는 복이배체(2n = 38, AACC) 식물로 전초를 나물용이나 가축의 청예사료로 이용하고, 유채기름은 튀김이나 샐러드와 같은 식용으로 이용되며, 바이오디젤의 원료로 이용된다. 착유 후 부산물인 유채박은 가축사료와 유기질 비료로도 사용되는 등 다양하게 이용하고 있다(Lee et al., 2015). 최근에는 우리나라에서도 대규모 유채 경관단지 조성을 통하여 지역 자치단체의 홍보와 함께 관광수익을 올리기 위해 남부지방 뿐만 아니라 중부지방에서도 경관을 목적으로 유채 재배면적이 증가하고 있다(Kim et al, 2015). 유채는 경관용 작물로서 관심의 대상이 되고 있으나 유채의 꽃색은 노란색으로 단순하여 경관효과를 높이기 위해서 꽃색을 다양화할 필요가 있다. 종간교배 및 속간교배를 통한 잡종 식물체의 생산은 불화합성 등의 이유로 어렵고 성공할 확률이 낮다(Myers, 2006). 그렇지만 유채의 육종은 양배추(B. oleracea L.), 겨자(B. juncea var. crispifolia L. H. Bailey) 및 배추(B. campestris ssp. pekinensis) 등의 십자화과(Cruciferae) 작물들과의 종간교배를 통해 유용한 형질의 도입에 대한 연구가 진행되었을 뿐만 아니라(Bajaj et al, 1986; Zhang et al, 2004), 무(Raphanus sativus L.)나 소래풀[Orychophragmus violaceus (L.) O. E. Schulz]과 같이 십자화과의 다른 속의 작물과의 속간교배에 대한 연구도 많이 진행되었다(Lelivelt et al, 1993; Li et al, 1995; Myers, 2006). 또한 속간 인공교배나 세포융합에 의해 꽃잎의 색이 붉은색이나 흰색으로 변화된 잡종 식물체도 보고된 바 있다(Hu et al, 2002; Wang et al, 2006; Zhao et al, 2008). 국내에서는 십자화과 Brassica 속 작물 사이의 종간교배에 대한 연구가 이루어지고 있으나(Kim et al, 2012; Lee et al, 2015; Seo et al, 2017), 속간교배에 대한 연구는 이루지지 않고 있다. 유채와 같은 십자화과에 속하는 양배추는 꽃이 크고 미색 또는 하얀색의 꽃이 피며, 제초제에 대한 저항성과 내병성이 있으며, 식물의 불화합성을 조절하는 복대립유전자가 있어 유채와 종간교배의 육종재료로 사용되고 있다(Ayotte et al, 1987). 또한 무는 흰색과 연보라색의 꽃이 피고(Wang et al, 2006), 소래풀은 ‘Chinese Violet Cress’ 또는 ‘February Orchid’로 불리며 자주색이나 분홍색 꽃이 피어 유채의 꽃색 육종에 있어서 유용한 유전재료로 이용이 가능하다(Luo et al, 1994). 본 연구에서는 유채와 같은 십자화과에 속하지만 다른 꽃색을 가진 양배추, 무, 소래풀 등의 유전형질을 유채에 도입하여 다양한 꽃색을 갖는 유채의 육종을 위한 기초 자료를 확보하기 위하여 종·속간교배를 실시하고, 종속간 인공교배에 따른 교배효율과 교배결과 만들어진 F1 잡종 식물체의 꽃색 및 꽃의 크기 등 식물학적 특성 및 지방산조성 등을 조사하였다.
재료 및 방법
시험재료
본 연구에 종자친으로 사용한 시험재료 중 유채(B. napus)는 농촌진흥청 국립식량과학원 바이오에너지작물연구소에서 육성한 ‘영산유채’, ‘탐미유채’(Jang et al, 1997), ‘한라유채(Rho et al, 1986)’ 3품종을 이용하였다. 화분친으로는 농촌진흥청 농업유전자원센터로부터 분양 받은 양배추(B. oleracea ‘VI033139’), 무(R. sativus ‘VI037218’), 보라색 꽃이 피는 소래풀[O. violaceus ‘violet’(가칭)] 을 사용하였다(Fig. 1).

인공교배
유채, 양배추, 무 및 소래풀 종자를 2013년 10월에 배양용 상토(바이오상토 1호, 흥농씨앗, 대한민국)를 채운 50구 육묘상자(4.5 × 4.5 × 5.0 ㎝)에 파종하고 본엽 2~3매 시기에 10℃에서 2주간 저온 처리하고, 본엽이 5매 정도 전개된 식물체를 20~25℃/10℃ (주/야간온도)로 조절되는 온실에 정식하여 자연일장 하에서 재배하였다. 인공교배는 개화 직전의 꽃봉오리를 핀셋을 이용하여 꽃잎과 수술을 제거하고, 다른 꽃가루의 수분을 방지하기 위하여 유산지 봉투로 밀봉하였다. 다음날 오전 9~11시 사이에 화분친의 꽃을 이용하여 인공수분 시키고, 다시 유산지 봉투로 밀봉한 다음 꼬투리가 완전하게 형성되는 10일 전후에 유산지 봉투를 벗기고 50~60일 후 수확하여 종속간 교배효율을 조사하였다.
종간교배효율 및 F1 식물체 표현형질 조사
종간 및 속간 교배효율은 인공수분 10일 후 꼬투리 형성여부를 조사 하였고, 50~60일 후에 꼬투리 형성율 및 꼬투리 당 결실 종자수 등을 조사하였다. 종속간 잡종 식물체의 표현형질 특성을 조사하기 위하여 결실 후 수확한 종자를 저온처리하지 않고 바로 배양용 상토를 채운 50구 육묘상자에 파종하고 25℃에서 발아시킨 후에 본엽이 2매 정도 전개된 시기에 10℃에 2주간 저온 처리하고, 본엽이 4매 정도 전개되었을 때 배양용 상토로 채운 화분에 이식하였다. 잡종식물체는 재배과정 중 하고현상을 줄이고 병충해를 방지하고자 약 2 m 높이에 차광용 검은색 차광막(55%)를 치고 통풍이 잘되는 곳에서 재배하였으며, 재배 30일 후 F1 잡종 식물체의 발아율, 외부형태 등의 농업형질을 조사하였으며, 개화 후 꽃색 및 꽃의 크기 등을 조사하였다.
지방산조성 분석
F1 잡종식물체 종자의 지방산 분석은 Kim et al (2013)의 방법에 따라 0.2 g의 시료를 막자사발에 분쇄한 후, 50 ㎖ 플라스크에 넣어 15 ㎖의 methyl alcohol과 1 ㎖의 sodium methoxide 30 wt%을 혼합하여 100℃ 온수조에서 2시간 동안 메틸화반응을 실시하였다. 반응물을 상온에서 1시간 동안 냉각시키고, 95% n-hexane 15 ㎖을 첨가하여 진탕 혼합한 후, 상층액을 1 ㎖를 취하여 0.2 ㎛ syringe filter에 여과하고 gas chromatography (Agilent 7890A, USA)로 지방산 조성을 분석하였다. 분석컬럼은 HP-INNOWAX (Agilent Co., 19091N-113, 30 m × 0.32 ㎜, 0.25 ㎛) silica capillary column을 사용하였다. 검출기의 온도는 주입부는 200℃, flame ionization detector (FID)는 250℃로 사용하였다. Oven온도는 140℃에서 10분간 유지한 다음 250℃에 도달하기까지 분당 6℃씩 상승시켰으며 250℃에서 5분간 유지, 각 지방산이 머문 시간을 확인하여 각 peak의 면적을 상대적인 백분율로 나타냈다.
결과 및 고찰
종속간 교배효율
Table 1에 유채와 십자화과 작물을 이용하여 인공적으로 종속간 교배를 실시하고 꼬투리 형성율과 꼬투리 당 종자 형성율을 나타내었다. 유채, 양배추, 무 및 소래풀을 온실에서 재배하면서 개화기에 인공수분을 실시하여 형성된 꼬투리의 수와 꼬투리 당 형성 종자의 수를 조사하였다. 유채와 양배추의 종간교배는 총 637개의 꽃을 인공수분하여 형성된 꼬투리는 419개로 65.8%의 꼬투리 형성율을 보였으나, 형성된 꼬투리 중 종자가 형성된 꼬투리 수는 23개였으며, 총 52개의 F1 잡종 종자가 생성되어 꼬투리 당 평균 종자수(seeds/set siliques)는 0.12개로 나타났다. 양배추를 화분친으로 이용하였을 때 유채 품종에 따른 꼬투리 형성율은 ‘영산유채’ × 양배추 ‘VI033193’ 교배조합이 100%로 가장 높게 나타났으며, 다음으로 ‘탐미유채’ × 양배추 ‘VI033193’ 66.4%, ‘한라유채’ × 양배추 ‘VI033193’ 58.7%, 순이었다. 꼬투리 당 형성된 종자 수는 ‘탐미유채’ × 양배추 ‘VI033193’의 조합이 0.17개로 가장 많았고, ‘한라유채’ × 양배추 ‘VI033193’ 교배조합에서는 꼬투리 당 0.13개의 종자가 생산되었다. ‘영산유채’ × 양배추 ‘VI033193’의 조합에서는 인공수분한 꽃에서 전부 꼬투리가 형성되었지만, 꼬투리 당 형성된 종자 수는 0.03개로 가장 적게 나타났다(Table 1). 십자화과 식물의 종간 교배에 대한 연구가 많이 진행되었는데, 유채와 양배추 종간 인공교배되었을 때 꼬투리 형성율이 22.8%였으며 꼬투리 당 종자 수는 0.12개로 보고하였고, Chiang et al (1977)은 유채와 배추를 교배하였을 때 꼬투리 형성율과 꼬투리 당 종자 수가 각각 90.6%, 15.5개, 유채와 순무(B. rapa L.)를 교배하였을 때 67.3%, 11.6개로 보고하였다(Kim et al, 2015). 이와 같이 동일한 Brassica 속에 속하는 식물사이의 인공교배에서도 꼬투리 형성율 및 결실 종자수에서 차이가 크게 나타나는 것은, 종간교배를 통한 잡종생산 시 종자친과 화분친의 유전형에 교배화합성에 차이가 있음을 나타낸 것으로 생각된다.
Table 1. The silique set and seed set from interspecific and lntergeneric cross of B. napus and B. oleracea., B. napus and R. sativus, B. napus and O. violaceus |
|
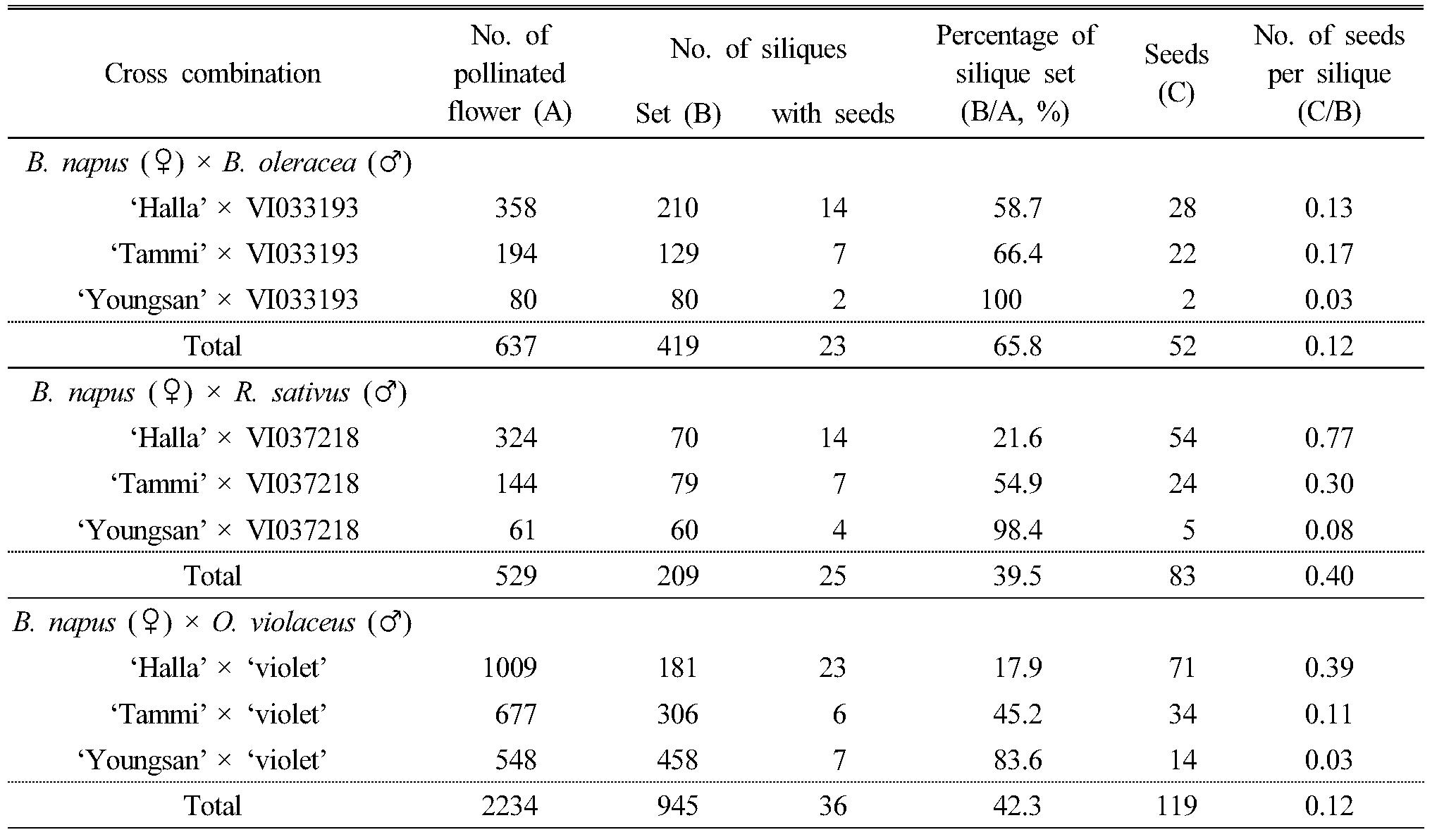
유채와 무의 속간교배는 총 529개를 인공수분하여 형성된 꼬투리 수, 결실된 꼬투리 수 및 종자 수를 조사한 결과, 총 209개의 꼬투리가 형성되어 39.5%의 형성율을 나타냈으며, 그 중 종자를 가진 꼬투리의 수는 25개였으며, 총 83개의 F1 잡종 종자를 얻어, 꼬투리 당 평균 종자 수는 0.40개로 나타났다. 유채 품종에 따른 교배효율은 ‘한라유채’를 종자친으로 사용하였을 때 324개를 교배하여 70개의 꼬투리가 형성되어 21.6%의 형성율을 보였고, 54개의 F1 잡종 종자를 얻어 꼬투리 당 종자 수가 0.77개로 가장 많았다. ‘탐미유채’를 종자친으로 144개를 교배하여 형성된 꼬투리의 수는 79개로 54.9%의 형성율을 보였고 종자가 형성된 꼬투리 수는 7개로 24개의 F1 잡종 종자를 얻어 꼬투리 당 종자 수는 0.30개였다. ‘영산유채’는 61개를 교배하여 형성된 꼬투리는 60개의 꼬투리를 얻었지만 그 중 종자를 가진 꼬투리는 4개로 5개의 F1 잡종 종자를 얻었으며 꼬투리 당 종자 수는 유채 품종 중 가장 적은 0.08개였다(Table 1). Lelivelt et al (1993)은 유채와 무의 속간교배를 통해 24.1%의 꼬투리형성율과 0.24개 꼬투리 당 종자수를 보고하여, 교배 시 유전형에 따라 꼬투리 형성율과 꼬투리 당 종자 수 등에서 차이가 있음을 확인하였다.
유채와 소래풀의 속간교배는 총 2,234개를 인공수분한 결과, 42.3%인 945개의 꼬투리가 형성되었으며 그 중 종자를 가진 꼬투리는 36개였고, 총 119개의 F1 잡종 종자를 얻어 꼬투리 당 평균 종자 수는 0.12개로 나타났다. 종자친으로 사용한 유채 품종에 따른 꼬투리 형성율과 꼬투리 당 평균 종자수는 각각 ‘한라유채’가 17.9%, 0.39개, ‘탐미유채’는 45.2%, 0.11, ‘영산유채’는 83.6%, 0.03개였다(Table 1). Cheng et al (2002)은 유채와 소래풀의 속간교배를 통해 꼬투리 당 종자수는 0.18개로 보고하였고, 꼬투리의 결실율 및 꼬투리당 평균 종자수는 종자친과 화분친의 유전형에 따라 달라진다고 보고하여 본 연구의 결과와 유사하였다. 속간교배는 종간교배보다 종간 유연관계가 멀기 때문에 성공률이 낮다고 알려져 있었지만, 본 실험에서는 꼬투리 형성율은 유채와 양배추의 종간교배에서 65.8%로 가장 높았고, 유채와 소래풀 속간 교배조합이 42.3%, 유채와 무의 속간교배조합이 39.5%순이었다. 꼬투리 당 형성된 종자 수는 유채와 무의 속간 교배조합에서 0.40개로 가장 높았으며, 유채와 양배추의 종간교배와 유채와 소래풀의 속간교배의 꼬투리 당 종자 수는 0.12개로 비슷하였다.
인공수분 후 형성된 꼬투리의 모양도 교배조합에 따라 달랐는데 유채와 유채를 교배하였을 때는 꼬투리의 모양이 반듯하며 길이가 5~6 ㎝ 정도로 길고 꼬투리 안에 빈곳이 없이 종자가 들어있는데 반해 종간 교배나 속간교배를 실시하면 인공수분 후 주두 부분과 꼬투리로 발달할 자방 부위가 노랗게 변하면서 꼬투리가 줄기로부터 떨어지거나, 꼬투리가 발달한 경우에도 길이가 매우 짧아지면서 꼬투리 모양이 불규칙하게 생장하였으며 이 경우 대부분 종자가 생성되지 않았다(Fig. 2).

잡종1세대의 식물학적 특성
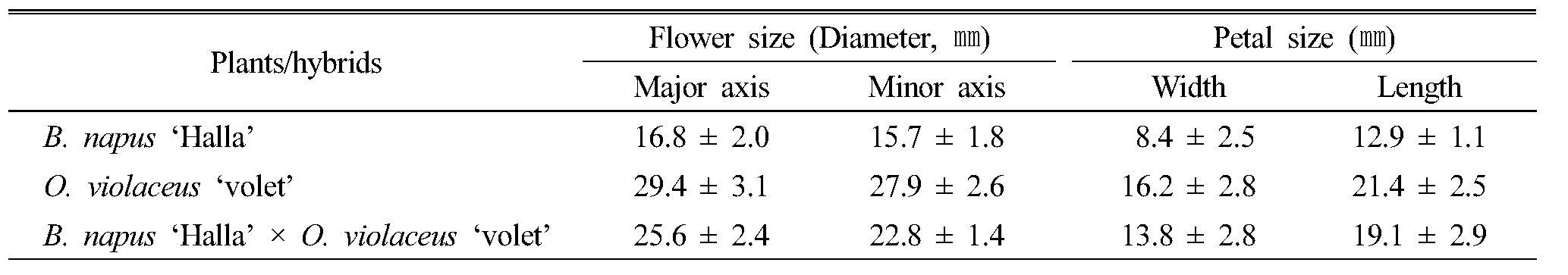
종속간 인공교배를 통해 획득한 잡종 식물체의 표현형질 특성을 조사하기 위하여, 인공수분 후 수확한 종자를 육묘판에 파종하고 25℃에서 발아시켜, 본엽이 4매 정도 전개되었을 때 화분에 이식하여 재배하면서 식물학적 특성을 관찰하였다. 종속간 교배를 통해 만들어진 F1 잡종 종자의 발아율은 67~92%로 종자친과 화분친 종자의 발아율 95% 이상에 비해 낮게 나타났다. 유채와 양배추의 종간교배 F1 잡종식물체의 잎 모양은 장란형이고 잎 표면과 뒷면에 모용이 없으며, 전열 형태의 결각을 나타내 전반적인 외부 형태는 종자친인 유채와 거의 유사하였고, 잎색은 회녹색을 띄어 있어 화분친인 양배추와 비슷하였다. 유채와 무사이의 속간교잡 F1 잡종식물체의 전반적인 잎의 형태도 종자친인 유채와 유사하였으나, 잎 가장자리에 유채는 거치가 없는데 반해 치아상의 거치가 발달하고와 잎 표면에 모용이 다수 발생하였고 잎자루가 유채나 무에 비하여 길어지는 특징을 나타냈다. 유채와 소래풀의 속간교잡 F1 잡종식물체도 외부형태는 종자친인 유채와 유사하였으나, 잎 표면에 모용이 발생하였고 잎자루가 길며, 엽의 폭이 넓어지는 특징을 나타냈다(Fig. 3, Table 2). Sabharwal and Dolezel (1993)은 유채와 배추를 정역교배(reciprocal cross)를 실시하여 유채를 모본으로 한 F1 잡종 식물체의 잎의 모양, 크기와 색이 유채와 유사하다고 보고하였으며, 배추를 모본으로 한 F1 잡종 식물체의 식물학적 특성 또한 유채와 유사하다고 보고하였다. Chiang et al (1977)의 유채와 양배추의 교잡 연구, Zao et al (2008)의 유채와 소래풀의 속간 교잡, Li et al (1998)은 갓(B. juncea)와 소래풀 및 Chengzhen et al (2014)의 배추(B. campestris)와 무의 교잡에서 생산된 F1 잡종의 형태적인 표현형이 양친의 중간형이거나 유채에 쪽에 좀 더 가까운 외부 형태를 보인다고 하였다. 하지만 Li et al (1995)의 연구에서는 유채와 소래풀의 F1 잡종 식물체의 특성은 잎이 짙은 녹색을 띄며 털이 많은 원형이며, 줄기도 분지된 가지 수가 많아 외부형태가 부계인 소래풀과 유사하다고 보고한 결과도 있다. 유채의 경관 효과를 높이기 위해서는 꽃수와 꽃색이 중요한데 유채와 종속간 교배를 통해 만들어진 F1 식물체의 꽃수는 화분친 보다 종자친인 유채의 꽃수와 비슷하였다. F1 잡종 식물체의 꽃색은 대부분 유채의 노란색 꽃색을 나타냈지만, ‘탐미유채’와 양배추 ‘VI033139’의 F1 잡종 식물체에서 유채의 노란색과 양배추의 흰색의 중간인 미색 꽃이 핀 개체가 출현하였다(Fig. 4). 또한 ‘한라유채’와 소래풀 ‘violet’의 속간교배 F1 잡종 식물체에서 꽃의 크기가 매우 큰 개체가 출현하였는데, 한라유채 꽃의 지름이 장축 16.8 ㎜, 단축 15.7 ㎜인데 반해 잡종식물체의 꽃의 크기가 각각 25.6 ㎜와 22.8 ㎜로 커졌으며, 이는 꽃이 큰 화분친인 소래풀(장축 29.4 ㎜, 단축 27.9 ㎜)의 형질이 발현되어 양친의 중간형을 나타낸 것으로 생각된다(Table 3). 유채의 노란색 꽃색은 꽃잎에서 합성되는 카로티노이드에 의해 결정되는데 흰꽃이 피는 양배추와의 교배에 의해 합성되는 카로티노이드의 양이 적어지면서 미색꽃이 피는 것으로 생각되며, 앞으로 F1 잡종 식물체를 대상으로 지속적인 여교배(backcross)를 통해 새로운 색을 갖는 유채품종을 만들어 낼 수 있을 것으로 생각된다.

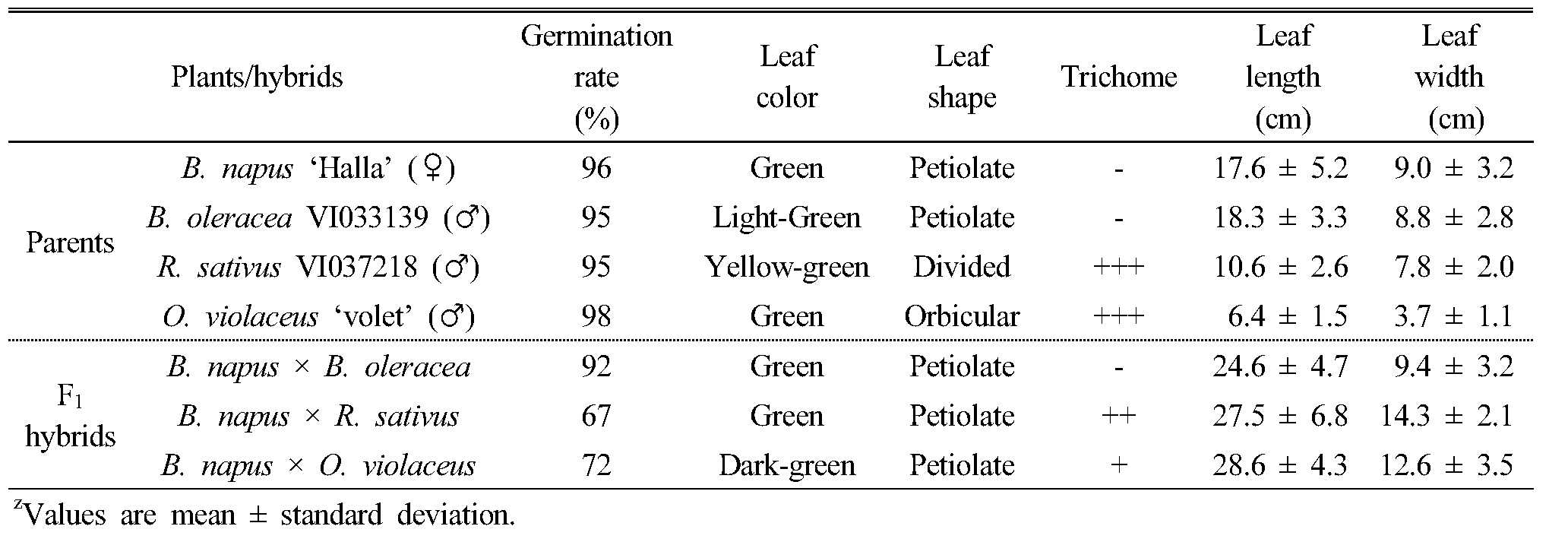

Table 3. Comparison flower and petal size of B. napus ‘Halla’, O. violaceus ‘volet’ and F1 hybrid B. napus ‘Halla’ × O. violaceus ‘violet’ |
|
지방산 조성
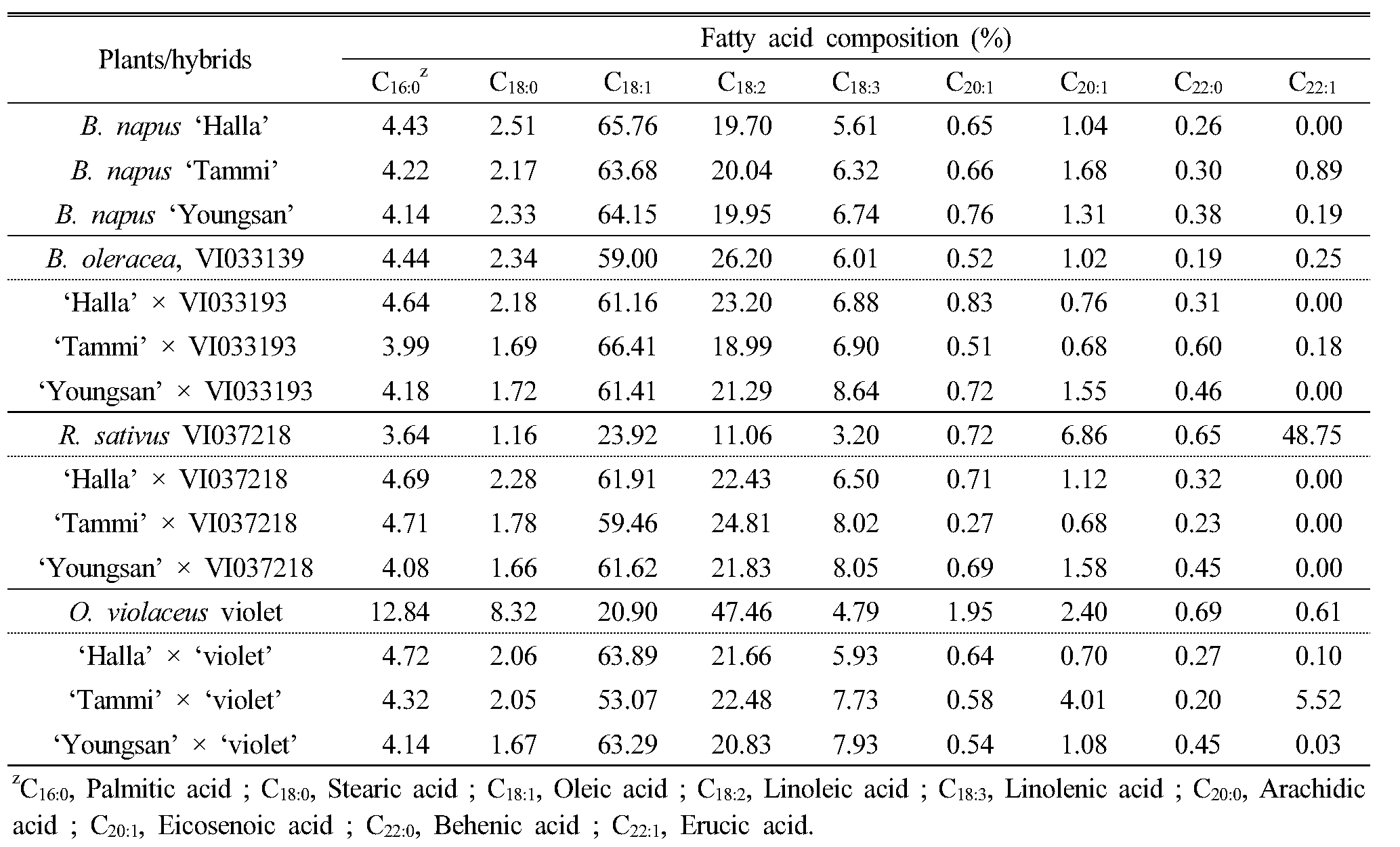
유채는 경관 감상용으로도 이용이 되지만 기름을 식용으로 이용하는 작물이다. 따라서 유채 종자 내의 기름중 지방산 성분도 중요한 육종 목표 중의 하나이다. 대부분의 십자화과 식물은 종자 내에 식용으로 부적합한 에루진산(erucic acid)이 다량 함유되어 있고, 이를 다량 섭취하였을 때 심장질환을 일으킬 수 있다는 연구 결과가 있다(Heijenskjöld and Ernster, 1975). 따라서 에루진산의 함량이 낮은 종·속간 잡종 식물체를 선발하기 위하여 종·속간교배를 통해 획득한 F1 잡종종자의 지방산 조성을 gas chromatography를 이용하여 분석하였다(Table 4). ‘한라유채’의 주요 지방산은 올레인산과 리놀레산이었으며, 각각 65.76%와 19.70%였고 에루진산은 검출되지 않았다. 반면에 양배추의 지방산 조성은 올레인산이 59.00%로 유채보다 낮았고 리놀레산이 26.20%로 유채보다 높았다. 유채와 양배추 F1 종자의 올레인산과 리놀레산의 함량이 각각 61.16%, 23.20%로 나타나 모계인 유채와 지방산 조성이 유사하였다. 무 종자의 지방산 조성은 올레인산 함량이 23.92%로 낮았고, 에루진산 함량이 48.75%로 매우 높게 나타났으나 유채와 무 속간교배 F1 종자의 지방산은 올레인산이 59~61% 정도 함유되었고, 에루진산은 전혀 검출되지 않아 모계인 유채와 유사한 형태의 지방산조성을 보였다. 소래풀의 주요 지방산은 올레인산과 리놀레산이지만 유채와는 다르게 리놀레산의 함량이 47.46%로 매우 높게 나타났으며, 팔미트산과 스테아릭 산도 각각 12.84%와 8.32%로 유채보다 높았다. 유채와 소래풀 F1 종자의 지방산 조성은 올레인산이 53.07~63.89%였으며 리놀레산의 함량도 20.83~22.28% 수준으로 유채와 비슷하였다.
적요
우리나라에서 유채는 식용유 생산과 경관을 목적으로 매년 재배면적이 증가하고 있다. 하지만 유채 꽃의 색상은 노란색으로 매우 단순하기 때문에 경관효과를 높이기 위해서는 유채 꽃색의 다양화가 필요하다. 유채 꽃색을 다양화하기 위해 유채와 십자화과 식물을 대상으로 종·속간교배를 실시한 결과, 유채와 양배추 종간 인공교배에서 꼬투리 형성율이 65.8%로 비교적 높았으며, 유채와 무, 유채와 소래풀 등 속간 교배조합에서의 꼬투리 형성율은 각각 39.5%와 42.3%로 낮았다. 인공수분 후 꼬투리가 발달하는 시기에 이종간 교잡 불화합성 때문에 종자 결실율이 매우 낮으며 꼬투리 당 생산된 평균 종자수는 유채와 양배추 교배조합에서 0.12개였으며, 유채와 무는 0.40개, 유채와 소래풀은 0.12개였다. 종속간 교배에 의해 생산된 F1 잡종식물체의 외부 표현형질은 잎의 형태와 꽃의 색은 모계인 유채와 유사하였으나, 잎의 길이와 폭 등이 전반적으로 커진 것으로 나타났다. 유채와 양배추의 종간교배에서 꽃의 색이 양친의 중간인 미색으로 피는 개체가 나타났고, 유채와 소래풀의 속간교배에서는 꽃의 크기가 커진 개체가 생겨 이 개체들을 육종재료로 이용하면 경관효과를 높일 수 있을 것으로 생각된다. 대부분의 F1 잡종종자의 올레인산, 리놀레산 및 리놀렌산 등의 지방산 조성은 양친의 중간 정도였으나, 유채와 무의 F1 잡종종자의 지방산 중 에루진산은 화분친인 무에 비교하여 급격하게 감소하였다.
