

Introduction
Materials and Methods
Plant materials and planting standards
Agronomic trait survey
Data analysis
Results
Agronomic traits
Screening of germplasms suitable for mechanized harvesting
Correlation analysis results
Principal component analysis results
Discussion
Introduction
Adzuki beans (red bean, Vigna angularis) are planted widely in more than 30 countries but are grown and consumed mainly in Korea, China, and Japan. Adzuki beans are rich in nutrients and have a sweet taste, and are used as a raw material for traditional pastries in East Asian countries. In traditional medicine, adzuki bean extract (Pa-EE) has anti-inflammatory effects (Yu et al., 2011) and is used in Korea, China, and Japan to treat arthritis, enteritis, and nephritis (Lim et al., 2020). Adzuki bean polysaccharide has antioxidant and immunomodulatory functions (Yao et al., 2015), and can regulate blood sugar and lipid metabolism in diabetic patients (Wu et al., 2020). Adzuki beans contain antioxidants such as policosanol, γ-tocopherol, phenolic compounds, flavonoids, and catechin (Kim et al., 2020; Lee et al., 2021). However, the antioxidant content differs depending on the variety and growing region. Adzuki bean distilled water fractions affect hair pigmentation (Itoh and Furuichi, 2005). In addition, during processing, the maltol component of adzuki beans can produce a unique caramel aroma that can enrich the flavor of pastries (Bi et al., 2021).
Adzuki beans are essential in food culture and agricultural systems. They fertilize the soil by fixing nitrogen and are popular for intercropping and rotation systems (Wang et al., 2019). For example, adzuki beans are often intercropped with cotton and cereal crops in northern China and sugarcane and fruit trees in southern China and are often grown in rotation in Japan. However, adzuki bean yields vary greatly due to differences in cultivation intensity, environmental factors, and the mechanization level. High yields of 20 - 30 tons per hectare were observed in Japan and northern China as a result of advanced breeding research and mechanization (Wang et al., 2019). However, adzuki bean yields are much lower in most countries. Adzuki beans are cultivated mainly in small-scale farming systems, associated with high costs and low economic benefits. Thus, Chinese and Korean breeders have striven in recent decades to achieve high yields, upright plants, and early maturity to save labor costs and increase incomes (Wang et al., 2019). Mechanized harvesting has been achieved in Japan, and cultivars with high bottom pod heights (BPHs) and lodging resistance have been developed. Currently, cultivars with strong disease resistance, thin seed coats, and starch granules suitable for sweet bean paste are being bred (Wang et al., 2019). The genetic diversity among adzuki bean cultivars is low, and landrace and wild germplasms could be used as breeding materials and stored in major germplasm resource banks. The National Agrobiodiversity Center in South Korea preserves 3,000 adzuki bean germplasm resources, ranking second in the world after the 5,500 accessions of the Chinese Academy of Agricultural Sciences.
The BPH is an important trait, as it must meet the conditions for mechanized harvesting to ensure adequate harvests and reduce losses. The BPH also affects the quality of the bottom pod, as the pod tops can rot or cause seeds to germinate when they touch the ground under high humidity (Zdravković et al., 2005), or change their natural color under dry conditions. Based on the structure of mechanized harvesting systems, the BPH of adzuki beans must be > 10 ㎝. In Korea, research on adzuki beans focused on lodging resistance before the BPH was investigated. The “Seona” and “Hongjin” cultivars, created in 1999 by crossbreeding, had the advantages of lodging resistance and high yield, quality, and antioxidant activity (Song et al., 2018; 2019). The synchronous maturing of seeds is a disadvantageous trait in the natural state. Seeds mature at different times, helping to ensure that they spread at different times and in different environments. However, synchronous maturing is enormously significant for mechanized harvesting. Early harvesting may result in insufficient seed filling, lower yields, and excessive seed moisture content. Late harvesting can lead to pod cracking, reduced dry weight, tarnished seeds, and reduced quality. To realize mechanized harvesting, cultivars with high synchronous maturing are essential.
Most current cultivars were bred using traditional hybridization techniques. The whole genome sequencing of the adzuki bean genome was performed relatively late (in 2015), and only a few studies performed molecular marker-assisted breeding and low- resolution genetic mapping on the adzuki bean at the molecular level. Molecular breeding of the adzuki bean increased after its whole genome sequencing (Yang et al., 2015). In this study, 14 agronomic traits of 806 Korean adzuki bean landraces were evaluated to reveal important relationships between traits and provide information and breeding materials for further studies of breeding and production mechanisms.
Materials and Methods
Plant materials and planting standards
The seeds of 806 accessions of adzuki bean (Supplementary Dataset 1) were provided by the National Agrobiodiversity Center, National Institute of Agricultural Sciences, Rural Development Administration, the Republic of Korea (http://genebank.rda.go.kr, accessed on September 21, 2022). All adzuki bean germplasms were sown in Jeonju, Korea, on June 21, 2021, and harvested in October 2021. Ten plants of each germplasm were sown 20 ㎝ apart with a row spacing of 70 ㎝.
Agronomic trait survey
Data on ten quantitative traits (BPH, flowering date, maturity date, growth period, plant height, lodging score, synchronous maturing, 100-seed weight, number of pods per plant, and number of seeds per pod) and three qualitative traits (pod color, seed coat color, and growth habit) were recorded. We calculated the average yield per plant as the 100-seed weight × number of pods per plant × number of seeds per pod / 100. We determined that 714,300 plants could be cultivated on 10 hectares, based on a planting distance of 70 × 20 ㎝, and calculated the yield per 10 hectares.
Data analysis
We calculated descriptive statistics (minimums, maximums, means, standard deviations, coefficients of variation, skewness, and kurtosis), tested for group differences with ANOVA, used chi-squared and Fisher’s exact tests to test distributions, and used the KMO and Bartlett’s sphericity tests to test the suitability of the data with the trial version of XLSTAT (Microsoft Corporation, Charlotte, NC, USA). Clustering, principal component, and correlation analyses were performed using the “NbClust” (Charrad et al., 2014), “FactoMineR” (Lê et al., 2008), and “corrplot” (Wei et al., 2017) packages, respectively, in R v. 4.2.1 (Ihaka and Gentleman, 1996).
Results
Agronomic traits
Eleven quantitative traits (BPH, flowering date, maturity date, growth period, plant height, lodging score, synchronous maturing, 100-seed weight, number of pods per plant, number of seeds per pod, and yield) and three qualitative traits (pod color, seed coat color, and growth habit) of four Korean cultivars and 802 Korean landraces were recorded (Supplementary Dataset 1). BPHs > 10 ㎝ are known to be required for mechanized harvesting. Only 169 accessions had BPHs > 10 ㎝; 637 accessions had BPHs ≤ 10 ㎝. Except for the flowering date range, the other traits value ranges were lower in germplasms with BPHs > 10 ㎝ than in those with BPHs ≤ 10 ㎝ (Table 1). The significant characteristics of accessions with BPHs > 10 ㎝, determined by Duncan’s analysis, were earlier flowering, shorter plant heights, higher lodging resistance, lower synchronous maturing, fewer pods per plant, and lower yields. For the 806 accessions, the distributions of flowering and maturity date data were close to normal, and average values were 63.769 and 42.932 days, respectively. Compared with the normal distribution, the growth period data skewed to the left, indicating the presence of more early-maturing than late-maturing varieties. A high degree of kurtosis indicated a concentrated distribution of growth period data, with an average value of 106.571 days. The average lodging value was > 30%, and these data had a close-to- normal distribution. The number of pods per plant, number of seeds per pod, and yield data skewed to the right with high degrees of kurtosis.
Table 1.
Ten quantitative traits in accessions with bottom pod heights > 10 ㎝ and ≤10 ㎝.
| Traits | Value | Bottom pod height | ||
| ≤ 10 ㎝ | > 10 ㎝ | Total | ||
|
Flowering date (Days) | Range | 46.000-75.000 | 44.000-76.000 | 44.000-76.000 |
| Mean ± SD | 64.283±4.655 a | 61.834±5.476 b | 63.769±4.940 | |
| Variation coefficient | 0.072 | 0.089 | 0.077 | |
| Skewness | -0.365 | -0.198 | -0.393 | |
| Kurtosis | 0.477 | 0.474 | 0.542 | |
|
Maturity date (Days) | Range | 26.000-63.000 | 27.000-53.000 | 26.000-63.000 |
| Mean ± SD | 43.060±4.709 a | 42.450±5.194 a | 42.932±4.821 | |
| Variation coefficient | 0.109 | 0.122 | 0.112 | |
| Skewness | 0.017 | -0.725 | -0.191 | |
| Kurtosis | 0.966 | 0.414 | 0.912 | |
|
Growing date (Days) | Range | 82.000-125.000 | 82.000-116.000 | 82.000-125.000 |
| Mean ± SD | 107.209±4.682 a | 104.166±6.181 b | 106.571±5.184 | |
| Variation coefficient | 0.044 | 0.059 | 0.049 | |
| Skewness | -1.171 | -1.591 | -1.444 | |
| Kurtosis | 6.780 | 2.806 | 5.773 | |
|
Plant height (㎝) | Range | 38.667-103.667 | 39.000-95.000 | 38.667-103.667 |
| Mean ± SD | 60.796±10.165 a | 58.592±10.907 b | 60.376±10.357 | |
| Variation coefficient | 0.167 | 0.186 | 0.172 | |
| Skewness | 0.701 | 0.418 | 0.611 | |
| Kurtosis | 0.844 | 0.165 | 0.698 | |
| Lodging score | Range | 0-9 | 0-9 | 0-9 |
| Mean ± SD | 3.757±2.402 a | 3.233±2.504 b | 3.576±2.478 | |
| Variation coefficient | 0.639 | 0.775 | 0.431 | |
| Skewness | 0.674 | 0.597 | -1.313 | |
| Kurtosis | -0.101 | -0.431 | 0.355 | |
| Synchronous maturity | Range | 1-3 | 1-3 | 1-3 |
| Mean ± SD | 1.907±0.433 a | 1.775±0.519 b | 1.880±0.456 | |
| Variation coefficient | 0.227 | 0.292 | 0.242 | |
| Skewness | -0.488 | -0.230 | -0.461 | |
| Kurtosis | 1.879 | -0.145 | 1.242 | |
| 100 seed weight (g) | Range | 3.567-20.700 | 3.933-18.933 | 3.567-20.700 |
| Mean ± SD | 12.054±3.701 a | 11.868±3.203 a | 12.015±3.603 | |
| Variation coefficient | 0.307 | 0.270 | 0.3 | |
| Skewness | -0.041 | 0.025 | -0.025 | |
| Kurtosis | -0.911 | -0.532 | -0.839 | |
|
Number of pods per plant (Number) | Range | 10.333-136.000 | 8.667-102.000 | 8.667-136 |
| Mean ± SD | 42.534±17.648 a | 35.823±15.112 b | 41.127±17.363 | |
| Variation coefficient | 0.415 | 0.422 | 0.422 | |
| Skewness | 1.4 | 1.266 | 1.384 | |
| Kurtosis | 3.391 | 2.377 | 3.323 | |
|
Number of seeds per pod (Number) | Range | 3.800-14.000 | 4.000-9.100 | 3.800-14.000 |
| Mean ± SD | 7.046±1.073 a | 6.866±1.062 a | 7.008±1.073 | |
| Variation coefficient | 0.152 | 0.155 | 0.153 | |
| Skewness | 0.383 | -0.096 | 0.284 | |
| Kurtosis | 2.164 | -0.467 | 1.668 | |
|
Yield (t/10ha) | Range | 3.457-113.386 | 4.601-56.344 | 3.457-113.386 |
| Mean ± SD | 24.747±12.003 a | 20.598±10.179 b | 23.877±11.766 | |
| Variation coefficient | 0.485 | 0.494 | 0.493 | |
| Skewness | 2.026 | 0.898 | 1.869 | |
| Kurtosis | 7.867 | 0.581 | 7.19 | |
Wide differences were shown in three qualitative traits (Table 2). Among the 806 accessions, the pod color at harvest was mostly white (548 accessions), followed by light brown (154 accessions), black (54 accessions), and deep brown (45 accessions). The seed coat color sets were red (486 accessions), gray (150 accessions), black dots on red (72 accessions), white (34 accessions), black mottle on white (23 accessions), black (11 accessions), black dots on brown (6 accessions), green (6 accessions), and yellow (4 accessions). Most germplasms had an upright growth habit (761 accessions), followed by a medium upright habit (45 accessions). As we selected only cultivars and landraces, the sample included no climbing germplasm. We used chi-square and Fisher’s exact tests to ascertain the differences in the distributions of the qualitative traits in germplasms with BPHs ≤ 10 and > 10 ㎝ (Supplementary Dataset 1). No significant distributional difference between groups was found for the three qualitative traits tested.
Table 2.
Three qualitative traits in accessions with bottom pod heights > 10 ㎝ and ≤10 ㎝
Screening of germplasms suitable for mechanized harvesting
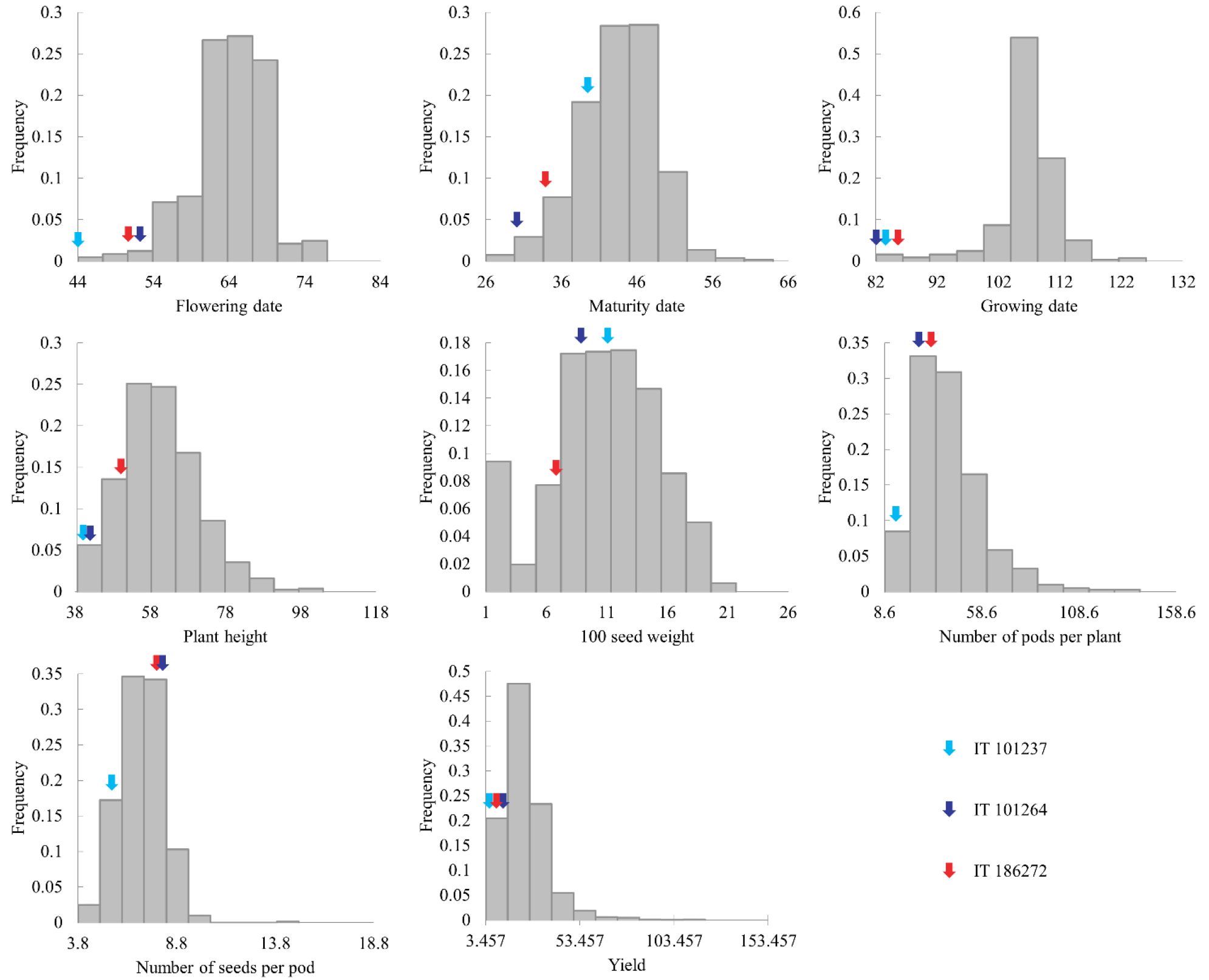
We screened accessions with BPHs > 10 ㎝, no lodging, and synchronous maturing rates > 76%. Three accessions met these criteria. They were all early flowering and maturing, with low plant heights and yields (Fig. 1). IT 101237 had the earliest flowering date (44 days after sowing) among all 806 accessions. IT 101264 and IT 186272 were also in the top 2% of early-flowering plants. IT 101264 matured earliest, with a growth period of 82 days; IT 101237 and IT 186272 had growth periods of 83 and 85 days, respectively, and the average growth period was 106.571 days. The plant heights of the three accessions (40.3, 41.0, and 51.3 ㎝, respectively) were below the average plant height (60.46 ㎝). The 100-seed weights of the three accessions were below the average (12.015 g). The 100-seed weight of IT 101237 was closest to the average at 10.5 g; those of IT 101264 and IT 186272 were 8.3 and 6.1 g, respectively. The number of pods per plant and yield of IT 101237, IT 101264, and IT 186272 were also below average.
Correlation analysis results
The selection of adaptive breeding materials suitable for mechanized harvesting revealed that the BPH, lodging score, and synchronous maturing potentially correlated with other traits. Thus, we conducted correlation analyses on 11 quantitative traits (Fig. 2) and performed ANOVAs to analyze differences between the qualitative and quantitative traits (Table 2) to reveal important associations between agronomic traits. Among the 11 quantitative traits, the number of pods per plant correlated strongly with the yield. Moderate correlations were detected among the three growth stage traits, and the BPH correlated weakly and negatively with the flowering date, growth period, synchronous maturing, lodging score, plant height, number of pods per plant, and yield. Synchronous maturity showed a weak positive correlation with the flowering date and a weak negative correlation with BPH. This negative correlation explains the reason that fewer germplasms were satisfying the conditions of high synchronous maturity and high BPH at the same time. The lodging score correlated weakly and positively with the flowering date, maturity date, and 100-seed weight and weakly and negatively with the BPH. A positive correlation between synchronous maturing and the flowering date was detected, with later flowering associated with more synchronous maturing.
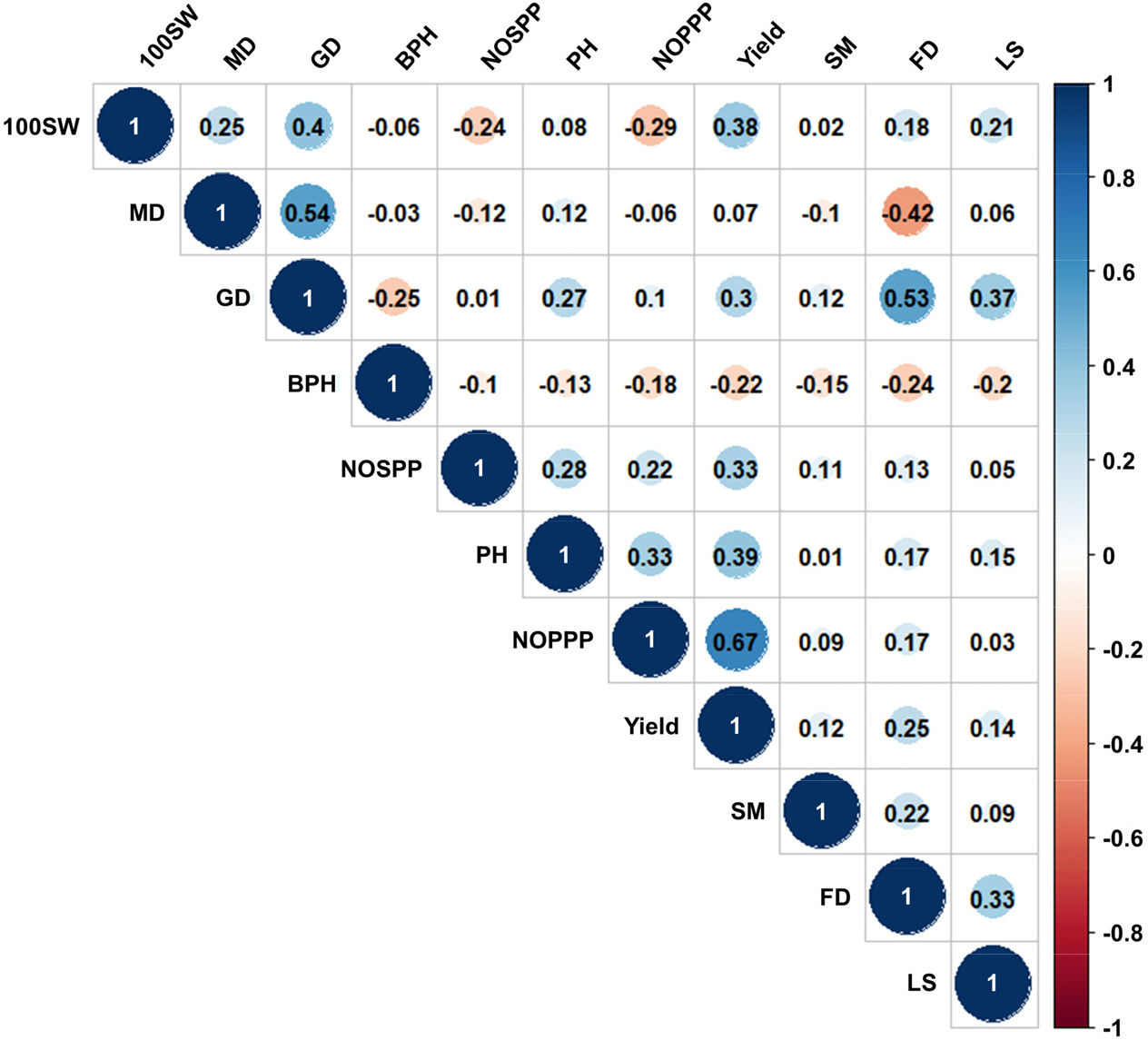
Fig. 2.
Correlation plot and coefficients for 11 quantitative traits. Blue represents a positive correlation and red represents a negative correlation. The circle size highlights the absolute value of correlation coefficient within the circle. 100SW, 100 seed weight; MD, Maturity date; GD, Growth date; BPH, Bottom pod height; NOSPP, Number of seeds per pod; PH, Plant height; NOPPP, Number of pods per plant; SM, Synchronous maturity; FD, Flowering date; LS, Lodging score.
Average times from flowering to maturity were 35.5 days for accessions with yellow seed coats and 43.9 days for those with black seed coats (difference, 8.4 days; Table S1). Accessions with brown seed coats with black dots had the greatest plant heights (average, 68.8 ㎝) and those with yellow seed coats were the shortest (53.0 ㎝; difference, 15.8 ㎝). The average lodging resistance score was 4.5-fold lower for accessions with brown seed coats with black dots than for those with yellow seed coats. The latter had no lodging, and average lodging rates for accessions with black seed coats were 11 - 50%. The 100-seed weight showed a significant difference; it was greatest for accessions with black seed coats (average, 16.812 g) and lowest for those with brown seed coats with black dots (average, 4.428 g; 3.8-fold difference).
The 100-seed weight, number of pods per plant, number of seeds per pod, and yield differed significantly among accessions with different pod colors (Table S2). The 100-seed weight was greatest for accessions with yellow-white pods (average, 12.868 g) and lowest for those with black pods (average, 9.381 g; difference, 3.487 g). Plants with black pods had the largest number of pods per plant (average, 47.019) and those with mixed-color pods had an average of only 31.333 pods per plant. Black pods also contained the largest number of seeds (average, 7.354) and white pods contained the fewest seeds (average, 6.898). The highest yield was for plants with white pods (average, 24.597 t/10 ha) and the lowest was for plants with mixed-color pods (average, 16.619 t/10 ha).
The population was divided into plants with upright (761 accessions) and intermediate (45 accessions) growth habits (Table S2). Because we selected only landraces and cultivars, the sample contained no climbing plants. The maturity date, growth period, plant height, 100-seed weight, number of pods per plant, and number of seeds per plant differed according to the growth habit. The average maturity dates for the intermediate and upright types were 104.792 and 106.661 days, respectively (difference, 1.869 days). The average difference in plant height was 14.791 ㎝, with averages for the upright and intermediate types of 59.550 and 74.341 ㎝, respectively. The average 100-seed weights for the upright and intermediate types were 12.294 and 7.293 g, respectively (difference, 5.001 g). The intermediate type had 58.289 pods per plant and the upright type had 40.112 pods per plant. The intermediate type also had more seeds per pod (7.720) than the upright type (6.966). None of the three qualitative traits correlated with any mechanization-adaptive trait (BPH, lodging score, or synchronous maturing; Tables S1 - S3).
Principal component analysis results
We performed principal component analysis (PCA) to evaluate the degree of variation in each trait, analyze the distributions of similar individuals, and reveal correlations between traits. The KMO test coefficient was > 0.5, indicating that the sample met the reasonable data structure requirement, and Bartlett’s test indicated that the data were independent (Supplementary Dataset 1). PCA of data on the 11 quantitative traits revealed seven components that explained 100% of the variance (Fig. 3). The first two principal components provided 25.3% and 19.5% resolution, respectively. Yield (20.04%) contributed most to PC1, followed by the growth period (18.18%), flowering period (13.49%), and plant height (11.29%); together, these traits provided > 60% of the contribution. The 100-seed weight (23.25%) contributed the most to PC2, followed by the maturity stage (21.98%), number of pods per plant (17.33%), number of seeds per pod (15.46%), and growth period (13.58%); together, these traits provided > 60% of the contribution. The “NbClust” package in R was used to determine that the optimal number of clusters was 3, and the k-means algorithm was used to divide the 806 accessions into three clusters based on data on eight quantitative traits. Germplasms with large numbers of pods per plant, large numbers of seeds per pod, and high yields were concentrated in cluster 1 (Fig. 4, Table S4). Germplasms with long growth periods and large 100-seed weights were concentrated in cluster 2. Finally, germplasms with high BPHs were concentrated in cluster 3. The PCA plot showed that the BPH correlated negatively with the lodging score and synchronous maturing in PC1 (Fig. 4d). On the PCA diagram divided into four regions along the coordinate axes, germplasms for which mechanized harvesting is suitable (according to the BPH) are located in the upper left and lower left areas. No lodging accessions are present in the upper left and lower left areas, and most germplasms with high synchronous maturing rates are in the upper right and lower right areas. This distribution explains why few germplasms met all three conditions. The distribution of traits showed that the BPH and synchronous maturing correlated negatively with the flowering date, maturity date, plant height, and lodging score, confirming the correlation analysis results.
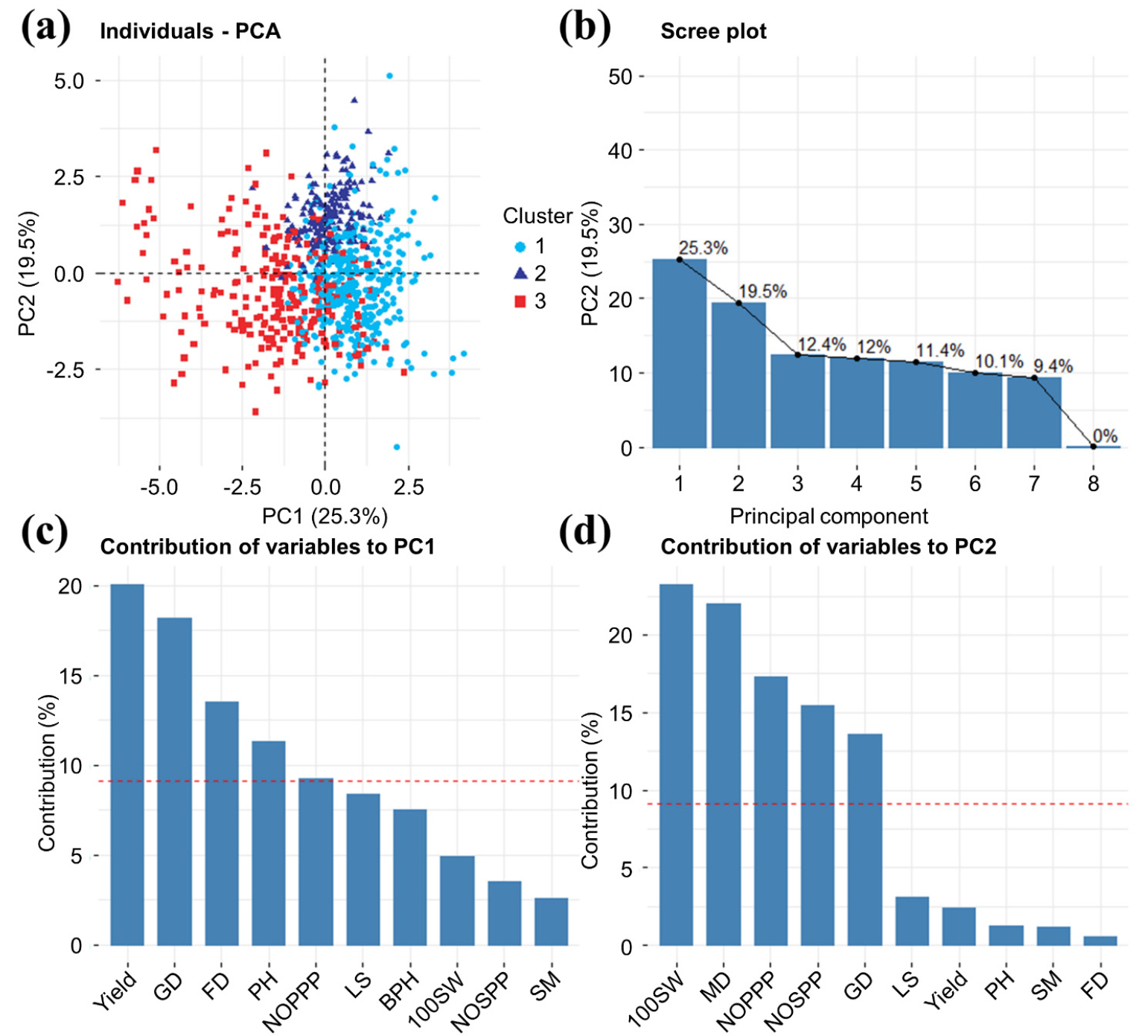
Fig. 3.
Distribution of 806 accessions according to 11 quantitative traits. (a) Three clusters were obtained by k-means clustering. (b) Discriminative power of PC1 - PC7. (c) Variable contributions to PC1. (d) Variable contributions to PC2. GD, growing date; FD, flowering date; PH, plant height; NOPPP, number of pods per plant; LS, lodging score; BPH, bottom pod height; 100SW, 100 seed weight; NOSPP, number of seeds per pod; SM, synchronous maturity; MD, maturity date.
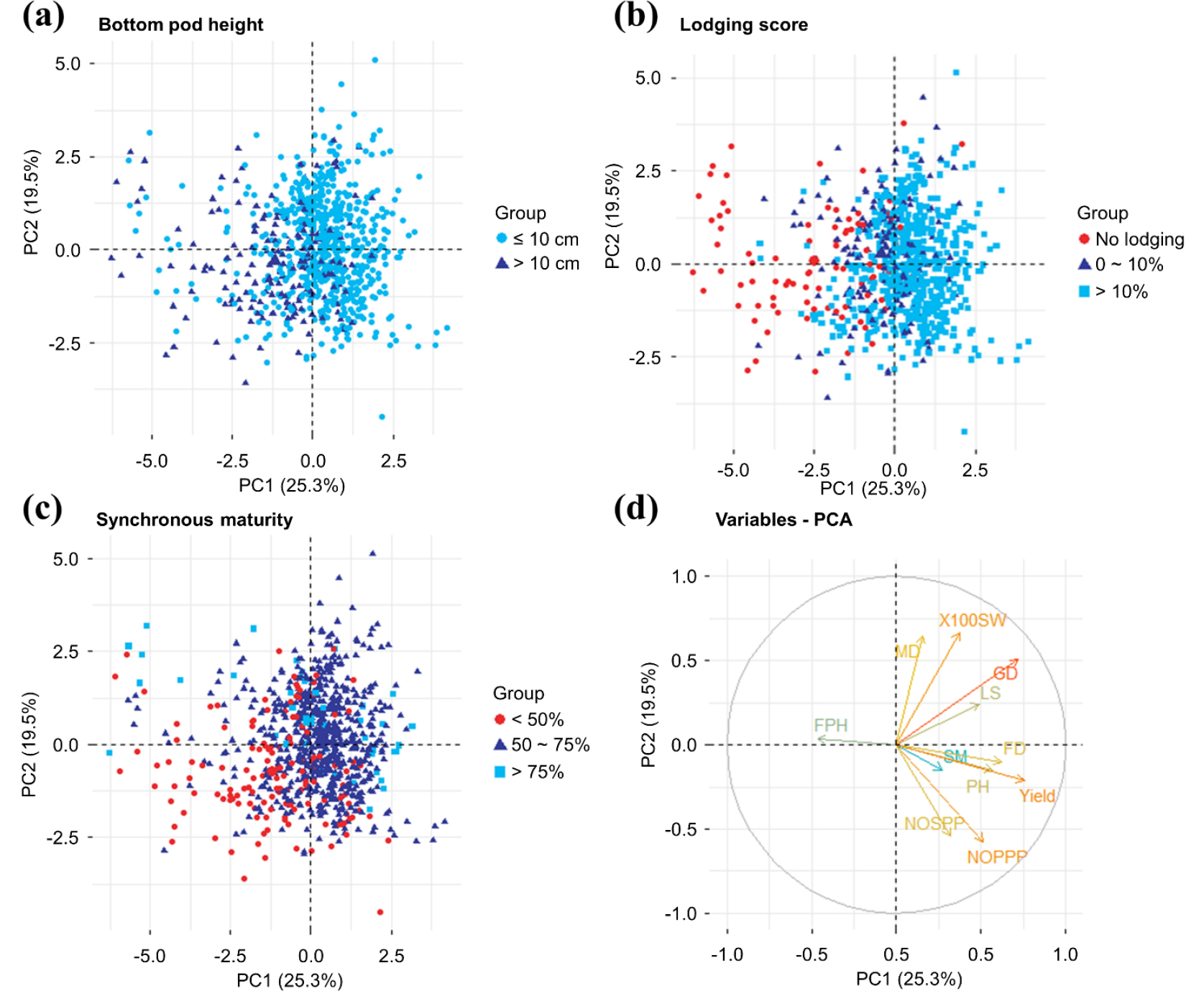
Fig. 4.
Principal component analysis of data on 11 quantitative traits in 806 accessions. (a) Colored by bottom pod height (BPH) ≤ 10 ㎝ and BPH > 10 ㎝. (b) Colored by lodging resistance. (c) Colored by maturity synchronization. (d) Distributions of 806 accessions according to 11 quantitative traits used to identify principal component 1 (24.3%) and principal component 2 (18.6%).
Discussion
Adzuki beans are used widely in desserts and traditional pastries because of their rich metabolites and unique flavor. Adzuki beans are cultivated mainly in traditional, small-scale farming systems, associated with high costs and low economic benefits. Thus, the development of cultivars that can be harvested by mechanized means is urgently needed. We analyzed 14 agronomic traits of adzuki beans to explore associations between adaptive and other traits with regard to mechanization suitability. Significant differences were observed in the agronomic traits of the 806 accessions. Yield showed the largest difference (CV = 0.493), followed by the lodging score (CV = 0.493) and number of pods per plant (CV = 0.422). The growth stage showed less difference, alongside the maturity date (CV = 0.112), flowering date (CV = 0.077), and growth period (CV = 0.049). We observed a strong positive correlation between the number of pods per plant and the yield, and this trait had the greatest impact on the yield. Consistent with this result, the number of pods per plant was previously identified as one of the most important factors affecting adzuki bean yields (Mirzaienasab and Mojaddam, 2014).
Moderate correlation among the three growth stage traits was observed, and the BPH, lodging score, and synchronous maturing all correlated weakly and negatively. Germplasms with high BPHs have low per-plant yields, reduced stress at the plant tops, and low plant heights, resulting in strong lodging resistance, which explains these findings. During the harvest period, a high yield may induce greater stress on the lodging resistance; however, the yield was related to the 100-seed weight, the number of pods per plant, and the number of seeds per pod. In addition, lodging resistance traits have been related to stem diameter and lignin content. The grain weight has been found to affect the lodging resistance of adzuki beans; large-grained, high-yielding, late-growing species are more prone to lodging (Song et al., 2018). Relatively comprehensive analyses of lodging resistance have led to the development of adzuki bean cultivars with excellent lodging resistance (Song et al., 2018; 2019).
Given the lack of previous studies of the BPH in adzuki beans, we compare our results with soybean BPHs. In this study, the BPH correlated weakly and negatively with the flowering date, growth period, synchronous maturing, lodging score, plant height, number of pods per plant, and yield. Consistent with these results, the soybean BPH has been found to correlate negatively with the number of pods, seeds per plant, and seeds per pod, and with the seed weight (Oz et al., 2009) and seed yield (Ghodrati et al., 2013). Consistent with our results, the BPH of soybean was found to correlate positively with the plant height (Oz et al., 2009). In addition, the BPH and plant height of soybean negatively affect the pod number and yield per plant, (correlation coefficient -0.322, p < 0.0001) (Kang et al., 2017), consistent with our findings. Thus, an ideal adzuki bean plant type must be developed to ensure that the yield with mechanized harvesting is suitable. Three germplasms with high synchronous maturing rates, BPHs, and lodging resistance were identified in this study. These three germplasms have the characteristics of early flowering and maturing, low plant heights, medium 100-seed weights, fewer pods per plant, and more seeds per pod. Thus, the 100-seed weight and number of pods per plant must be considered in breeding programs seeking to develop mechanization-adapted varieties to ensure good yields. Eight years of BPH data from 147 recombinant inbred soybean lines were used to identify quantitative trait loci; the findings suggested that eight auxin signaling-related genes are related to the BPH (Jiang et al., 2018). The auxin concentration negatively regulates soybean pod formation (Nonokawa et al., 2007) and internode elongation (Wilson et al., 1999). In addition, the soybean planting density has been found to correlate positively with plant height and BPH (Çalişkan et al., 2007; Gulluoglu et al., 2016). An increase in the planting density may increase the adzuki bean plant height and BPH, which are beneficial for mechanical harvesting. The combined use of high- yielding adzuki bean varieties and moderately increased planting densities may increase the suitability of mechanical harvesting, given the morphological characteristics of these varieties. This hypothesis will be tested in subsequent experiments.
To simulate the mechanized harvesting scenario, synchronous maturing is expressed as the proportion of individuals reaching the harvesting standard at the first harvest. Synchronous maturing correlated positively with the flowering date and negatively with the maturity date in this study. In other studies, synchronous maturing has been expressed as the time difference between the maturity of the first plant and the maturity of all plants. However, the correlation with other traits was consistent with our results. The best time to harvest is when 80% of the pods turn yellow and mature. The time from first pod maturity to this 80% standard varies among individuals, and we believe that this time difference impacts synchronous maturing. These traits will be examined in greater detail in our next planting, as we believe that the identification of links between them and the discovery of their key genes will enable the breeding of synchronously maturing varieties.
This study analyzed the widely differences in mechanized harvesting adaptive traits of adzuki bean resources, subdivided quantitative criteria, and revealed important correlations between these traits and other agronomic traits. Based on these correlations, we also propose a breeding scheme to improve the mechanization adaptability of adzuki bean, and the trait records from 806 accessions will be used to select the core collection. The core collection will be more effectively integrated with molecular techniques to facilitate crop improvement. Adzuki bean has undergone whole- genome sequencing and functional gene annotation (Yang et al., 2015). In future studies, suitable populations will be screened and re-sequencing to explore the relationships between and genetic backgrounds of these traits.
