

서 언
재료 및 방법
식물재료 및 ethephon, 염화칼슘 처리
과실 성숙도 판정
과실특성 분석
통계분석
결 과
Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과실 성숙도 변화
Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과피색 변화
Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과실특성 변화
고 찰
적 요
서 언
사과나무(Malus × domestica)는 장미과(Rosaceae) 낙엽교목 식물로, 우리나라 주요 과수작물 중 하나이다. 사과는 식이섬유와 비타민C 함량이 풍부하고 폴리페놀 화합물이 함유되어 있어 높은 항산화 활성을 나타낸다(Lee et al., 2018). 2020년 국내 사과 재배면적은 28,264 ha로 전체 과수 재배면적의 약 20%를 차지하고 있다(KOSIS, 2020). 최근 과원 갱신과 신규 과원 조성으로 사과 재배면적이 증가함에 따라 연간 사과 생산량 및 수요량도 더욱 증가할 것으로 예측되고 있다.
‘Fuji’ 사과는 10월 하순부터 수확되는 만생종으로 저장성이 상온에서 90일, 저온 저장시 150일 정도로 우수하여 우리나라에서 가장 많이 재배되고 있다. ‘Fuji’ 사과의 이론적인 수확시기는 만개 후 180일 전후로, 우리나라에서는 가을철 기상 상태에 따라 10월 중순부터 늦게는 12월 상순까지 수확을 한다(Sagong et al., 2013). 이와 같이 ‘Fuji’ 사과의 수확시기가 가변적인 이유는 생육기와 수확기 기온에 따른 과피 착색 때문이다. 사과는 생육기 기온이 높으면 ethylene 발생량이 높아 과실 성숙이 촉진되지만 과피 착색은 저조하다(Warringtone et al., 1999). 또한 ‘Fuji’ 사과의 수확기에 저온이 늦게 오면 과피 착색이 불량하여 착색 증진을 위해 수확시기를 연장할 수밖에 없다.
과피 착색에는 과실의 생리적 성숙 단계와 광, 온도, 지력과 수체 영양, 결실량, 스트레스, 유전인자 등 다양한 요인들이 관여한다(Marais et al., 2001). 사과 재배농가에서는 과피 착색 증진을 위해 하계 전정과 적엽, 반사필름 멀칭, 착색제를 사용하고 있으나 이는 경영비 상승의 주요 원인이다. 또한 대과 생산을 위해 과도하게 무기물을 투입하는 경우, 엽 내 질소 함량이 높아지고 착색이 지연되어 수확시기가 늦어진다. 이는 수체 내 저장양분 소비의 증가로 다음해 개화 및 결실 관리에 문제를 야기한다.
Ethephon (2-chloroethylphosphonic acid)은 성숙 및 노화와 관련된 식물호르몬인 ethylene 생성을 촉진하는 화합물로, 사과나무를 포함한 호흡급등형 과수작물의 당도, 산도, 경도, 착색 등 과실 성숙과 관련된 생리적 반응을 유도한다(Ban et al., 2009; Hayama et al., 2006). 식품의약품안전처 농약 잔류 허용기준에 따라 ethephon은 과수작물 중 사과, 체리, 포도나무에서 사용될 수 있도록 허가되어 있다.
포도 과방에 ethephon 분무처리는 수확시기를 2일 촉진하였으며(Kim and Chung, 2000), ‘Bing’ 체리(Prunus cerasus)에 수확 22일 전 ethephon 처리는 과피 착색을 27% 증진시키고 경도를 19% 감소시켰다(Smith and Whiting, 2010). 또한 ‘Fuji’ 사과의 수확 4주 전 100 ㎎/L ethephon 처리로 과피 착색이 증진되었음이 보고되었다(Li et al., 2002). 이와 같이 수확 전 ethephon 처리는 수확기를 촉진하고 과피 착색을 증진할 수 있으나, 세포벽 연화로 인해 과실 경도가 감소되는 문제가 발생한다(Dhillon and Mahajan, 2011; Hayama et al., 2006).
식물 생육의 필수 다량원소인 칼슘(Ca)은 세포 내 신호전달물질로, 세포의 기능을 조절하고 세포벽의 펙틴 중합체와 상호작용하여 세포벽 구조를 유지하는 역할을 한다(Palta, 1996; Singh et al., 2012). 염화칼슘(CaCl2), 키토산 등 칼슘이 포함된 화합물은 열과를 방지하고 병해 저항성을 증진하는 효과가 확인되어 다양한 원예작물의 안정적인 재배를 위해 활용되어 왔다(Kim et al., 2023; Michailidis et al., 2017). 따라서 사과 과실에 칼슘처리로 과실 경도 감소를 완화하고 저장 기간을 연장하여 식미가 우수한 사과를 유통할 수 있다(Ortiz et al., 2011).
‘Fuji’ 사과는 수확기에 저온이 늦게 오면 과피 착색이 불량하여 착색 증진을 위해 수확시기를 연장할 수밖에 없다. 하지만 수확 시기가 늦어지면 밀병현상이 발생하고 동결피해를 받을 수 있으며, 수확 후 장기저장 중 생리장해 발생이 증가할 수 있다(Sagong et al., 2013). 따라서 안정적으로 상품성이 높은 ‘Fuji’ 사과 생산을 위해 과실비대가 종료된 시점의 미숙과에 저농도의 ethephon 처리가 과피 착색 증진과 과실 성숙에 미치는 영향을 조사하였다. 또한, ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 ‘Fuji’ 사과의 과실 특성을 비교하여 안정적인 수확기 조절 기술을 제시하고자 하였다.
재료 및 방법
식물재료 및 ethephon, 염화칼슘 처리
본 시험은 충청북도 충주시 소재 사과원(36°58'7.61"N, 127°27'16.2"E)에서 수행하였다. 2020년에 수고 4.5 m의 7년생 ‘Fuji’/M.26 6주를 ethephon 무처리구(대조구) 3주와 ethephon 처리구 3주로 나누어 이용하였다. 2021년에는 2020년의 시험수와 생육이 유사한 7년생 ‘Fuji’/M.26 9주를 대조구 3주와 ethephon 처리구 3주, ethephon과 염화칼슘 혼합처리구 3주로 나누어 이용하였다.
과실 비대생장이 종료된 만개 후 145일에 100 ㎎/L의 ethephon을 수관 전체에 분무하였고, 대조구는 증류수를 분무하였다. 이후, 10일 주기로 대조구와 처리구에서 각각 4개의 과실을 무작위로 수집하여 성숙도 및 과실특성 분석에 이용하였다. 2020년에는 만개 후 185일까지 과실을 수집하였고, 2021년에는 만개 후 195일까지 과실을 수집하였다.
과실 성숙도 판정
대조구와 ethephon 처리구, ethephon 및 염화칼슘 혼합처리구에서 수집한 ‘Fuji’/M.26의 과실 성숙도 판정을 위해 과실 전분 반응지수를 조사하였다. 증류수 1 L에 10 g의 KI와 1.5 g의 I2를 용해시켜 제조한 요오드화칼륨 용액을 과실 횡단면에 스포이드로 도포 후 농촌진흥청의 ‘Fuji’ 사과 전분반응 지수를 참고하여 과실 성숙도를 조사하였다.
과실특성 분석
과중은 전자저울(AR3130, Ohaus Co., USA)로 측정하여 g으로 나타냈다. 과실 경도는 직경 5 ㎜의 plunger를 장착한 과실경도계(FHM-5, Takemura, Japan)를 이용하여 과실 적도면에서 임의로 3회 측정한 값을 평균하였다. 가용성고형물 함량은 당도계(Digital refractometer PR-101, Atago, Japan)를 이용하여 측정하였다. 적정산도 측정을 위해 사과 과육을 블렌더를 이용하여 분쇄 후 과즙을 수집하였다. 수집한 과즙에서 5 mL를 취해 증류수 20 mL과 혼합 후 10 mL을 추출하여 스포이드로 1% 페놀프탈레인 용액 2방울을 첨가하였다. 이 후, 0.1 N NaOH로 pH 8.3까지 적정하여 malic acid로 환산하였다. 과피색은 색차계(CR-300, Minolta, Japan)를 이용하여 hue 값을 측정하였고, 과실 적도면에서 임의로 5회 측정한 값을 평균하였다.
통계분석
대조구와 ethephon 처리구 간의 유의성을 검정하기 위해 R software v3.6.0을 이용하여 t-test를 수행하였다. 대조구, ethephon 단용처리구, ethephon 및 염화칼슘 혼합처리구 간의 유의성 검정은 R software v3.6.0을 이용하여 분산분석 후 5% 유의수준에서 던컨의 다중검정(Duncan’s multiple range test)을 수행하였다.
결 과
Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과실 성숙도 변화
‘Fuji’/M.26의 수확시기에 따른 전분함량은 ethephon 처리구와 ethephon 및 염화칼슘 혼합처리구에서 처리 10일 후(만개 후 155일)부터 감소하였다(Fig. 1). 2020년 만개 후 165일의 ethephon 단용처리구는 대조구보다 성숙이 촉진되었으며, 만개 후 175일의 전분함량과 유사하였다. 또한, 두 처리구에서 모두 만개 후 185일에는 밀병현상이 관찰되었다. 2021년 만개 후 175일에 ethephon 단용처리구의 전분함량은 만개 후 185일의 대조구보다 낮게 관찰되었으며, ethephon 및 염화칼슘 혼합처리구의 전분함량은 만개 후 185일과 유사하였다. 만개 후 195일의 전분함량은 세 처리구 중 대조구가 가장 높았으며, ethephon 및 염화칼슘 혼합처리구, ethephon 처리구 순으로 낮았다. 또한, 세 처리구 모두 밀병현상이 관찰되었다.

Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과피색 변화
Ethephon 처리에 따른 과피색 변화를 비교한 결과, 세 처리구 모두 만개 후 155일부터 과피 착색이 시작되었다(Fig. 2). 2020년의 ethephon 처리구는 만개 후 155일부터 과피 착색이 촉진되어 만개 후 165일에는 전면 착색이 완료되었다. 반면 대조구는 만개 후 175일에 과피 전면에 80% 이상 착색되었다(Fig. 2A). 2021년의 대조구는 ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구에 비해 과피 착색이 지연되는 것으로 관찰되었다(Fig. 2B). 만개 후 165일에는 대조구와 ethephon 및 염화칼슘 혼합처리구의 과피 착색율이 유사하였으며, ethephon 단용처리구는 상대적으로 착색이 촉진되었다. 만개 후 175일에 ethephon 단용처리구의 과피 착색은 만개 후 185일의 대조구와 ethephon 및 염화칼슘 처리구와 유사하였으며, 전면 착색이 완료되었다. 만개 후 195일에는 세 처리구에서 모두 전면 착색이 완료되어 육안상 과피색의 차이가 관찰되지 않았다(Fig. 2B).

과피의 hue값은 대조구, ethephon 단용처리구, ethephon 및 염화칼슘 혼합처리구에서 모두 만개 후 145일부터 지속적으로 감소하였다(Fig. 3). 2020년 만개 후 175일과 만개 후 185일에 두 처리구의 hue값은 유의한 차이가 없었다. 하지만 만개 후 165일에 ethephon 처리구의 hue값(42.0 ± 28.6°)은 대조구(93.4 ± 18.1°)보다 유의적으로 낮았다(p < 0.05). 또한 만개 후 165일에 ethephon 처리구의 hue값은 대조구의 만개 후 175일의 hue값(42.2 ± 8.3°)과 유사하였다(Fig. 3A).
2021년에는 만개 후 155일에 대조구(112.0 ± 2.1°)와 ethephon 처리구(102.9 ± 3.0°)의 hue값은 유의적인 차이를 나타냈지만 ethephon 및 염화칼슘 혼합처리구(110.5 ± 2.8°)의 hue값은 두 처리구와 유의적인 차이가 없었다. 만개 후 175일에는 과피 착색이 진행되며 hue값이 감소하였으나 ethephon 단용처리구(62.2 ± 5.1°)의 hue값이 대조구(83.1 ± 6.1°)보다 유의적으로 낮았다. 만개 후 185일에는 ethephon 처리구(49.9 ± 5.5°)와 ethephon 및 염화칼슘 처리구(43.7 ± 4.4°)의 hue값 모두 대조구(67.3 ± 5.2°)보다 유의적으로 낮았다. 또한, 만개 후 175일에 ethephon 및 염화칼슘 혼합처리구의 hue값은 만개 후 185일의 대조구의 hue값과 유사하였다(Fig. 3B).

Fig. 3.
Changes of hue values in control (CT), ethephon treated (ET), and ethephon and CaCl2 treated (EC) ‘Fuji’/M.26 fruits collected at different ripening stages in 2020 (A) and 2021 (B). DAFB, days after full bloom. The bars indicate ± SD (n = 4). Significant differences investigated by t-test are represent to * (p < 0.05), ** (p < 0.01), *** (p < 0.001), and ns (not significance). Different letters indicated significant differences between the treatments at p < 0.05.
Ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 따른 과실특성 변화
2020년 ‘Fuji’/M.26의 과실 비대생장 종료 후 수확시기에 따른 대조구와 ethephon 처리구의 과실특성 변화를 조사한 결과, 과중은 만개 후 145일에 278.6 ± 30.7 g으로 조사되었다. 대조구는 만개 후 165일에 339.7 ± 29.5 g이었으며, ethephon 처리구는 만개 후 175일에 365.8 ± 29.7 g으로 조사되었다. 두 처리구에서 모두 만개 후 185일까지 과중이 증가하는 경향을 보였으나 유의적인 차이는 없었다(Fig. 4A). 2021년에는 대조구와 ethephon 단용처리구, ethephon 및 염화칼슘 혼합처리구에서 모두 과실 비대생장 종료 후 만개 후 195일까지 과중이 증가하는 경향을 보였으나, 만개 후 185일을 제외하고 세 처리구의 과중간 유의적인 차이는 없었다. 만개 후 185일에는 ethephon 및 염화칼슘 혼합처리구의 과중(359.1 ± 35.9 g)이 ethephon 단용처리구의 과중(306.6 ± 25.1 g)보다 유의적으로 높았으나, 대조구(315.4 ± 21.1 g)와는 유의적인 차이는 없었다(Fig. 4B).

Fig. 4.
Changes of fruit weight in control (CT), ethephon treated (ET), and ethephon and CaCl2 treated (EC) ‘Fuji’/M.26 fruits collected at different ripening stages in 2020 (A) and 2021 (B). DAFB, days after full bloom. The bars indicate ± SD (n = 4). Significant differences investigated by t-test are represent to * (p < 0.05), ** (p < 0.01), *** (p < 0.001), and ns (not significance). Different letters indicated significant differences between the treatments at p < 0.05.
과실 경도는 2020년에 만개 후 165일까지 두 처리구 간의 유의적인 차이는 없었으나 만개 후 175일에 대조구의 경도는 0.76 ± 0.01 ㎏Ø5 ㎟, ethephon 처리구의 경도는 0.70 ± 0.01 ㎏Ø5 ㎟으로 ethephon 처리구의 경도가 유의적으로 낮았다(p < 0.01). 만개 후 185일에 두 처리구의 경도가 증가하는 경향을 보였지만, ethephon 처리구의 경도가 대조구보다 유의적으로 낮았다(p < 0.01, Fig. 5A). 2021년에는 수확 시기에 따라 세 처리구 간에 유의적인 차이는 없었다. 하지만 ethephon 단용처리구의 평균 과실 경도가 가장 낮았으며, ethephon 및 염화칼슘 혼합처리구의 평균 과실 경도는 만개 후 175일부터 195일까지 높은 수준을 유지하였다(Fig. 5B).

Fig. 5.
Changes of fruit firmness in control (CT), ethephon treated (ET), and ethephon and CaCl2 treated (EC) ‘Fuji’/M.26 fruits collected at different ripening stages in 2020 (A) and 2021 (B). DAFB, days after full bloom. The bars indicate ± SD (n = 4). Significant differences investigated by t-test are represent to * (p < 0.05), ** (p < 0.01), *** (p < 0.001), and ns (not significance). Different letters indicated significant differences between the treatments at p < 0.05.
당도는 2020년에 만개 후 145일부터 증가하는 경향을 보였으나 시기별 두 처리구의 유의적인 차이는 없었다(Fig. 6A). 2021년에도 만개 후 195일까지 증가하는 경향을 보였다. 만개 후 175일에는 ethephon 단용처리구의 당도(11.6 ± 0.5 °Brix)가 대조구(10.8 ± 0.4 °Brix)보다 유의적으로 높았으며, ethephon 및 염화칼슘 혼합처리구(11.5 ± 0.4 °Brix)의 당도는 두 처리구와 유의적인 차이가 없었다(Fig. 6B).
적정산도는 2020년 만개 후 175일에 급격히 감소하였다. 만개 후 175일에 대조구의 적정산도는 0.70 ± 0.04%, ethephon 처리구는 0.55 ± 0.11%로 유의적인 차이를 보였다(p< 0.05). 만개 후 185일에는 대조구의 적정산도가 감소하여 ethephon 처리구와 유의한 차이가 없었다(Fig. 6C). 2021년에는 세 처리구에서 모두 만개 후 145일 이후 감소하는 경향을 보였다. 만개 후 165일과 175일에 대조구는 적정산도가 유의적으로 높았으며, ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구는 유의적인 차이가 없었다. 만개 후 185일에는 세 처리구에서 0.46 ± 0.05~0.50 ± 0.07% 범위의 적정산도 값이 측정되었으며, 유의적인 차이가 없었다(Fig. 6D).
2020년에 조사된 당산비는 전 시기 동안 두 처리구 간 유의적인 차이는 없었으며, 만개 후 175일에 급증하는 경향을 나타내었다(Fig. 6E). 2021년에는 수확시기가 지남에 따라 증가하는 경향을 보였으며, 만개 후 165일과 175일에 ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구의 당산비는 대조구보다 유의적으로 높았다. 만개 후 185일과 195일에 세 처리구의 당산비는 유의적인 차이가 없었다. 또한 만개 후 175일의 ethephon 단용처리구(24.8 ± 1.0)와 ethephon 및 염화칼슘 혼합처리구(24.7 ± 2.5)의 평균 당산비는 만개 후 185일의 대조구(23.3 ± 2.9)의 당산비보다 높았다(Fig. 6F).
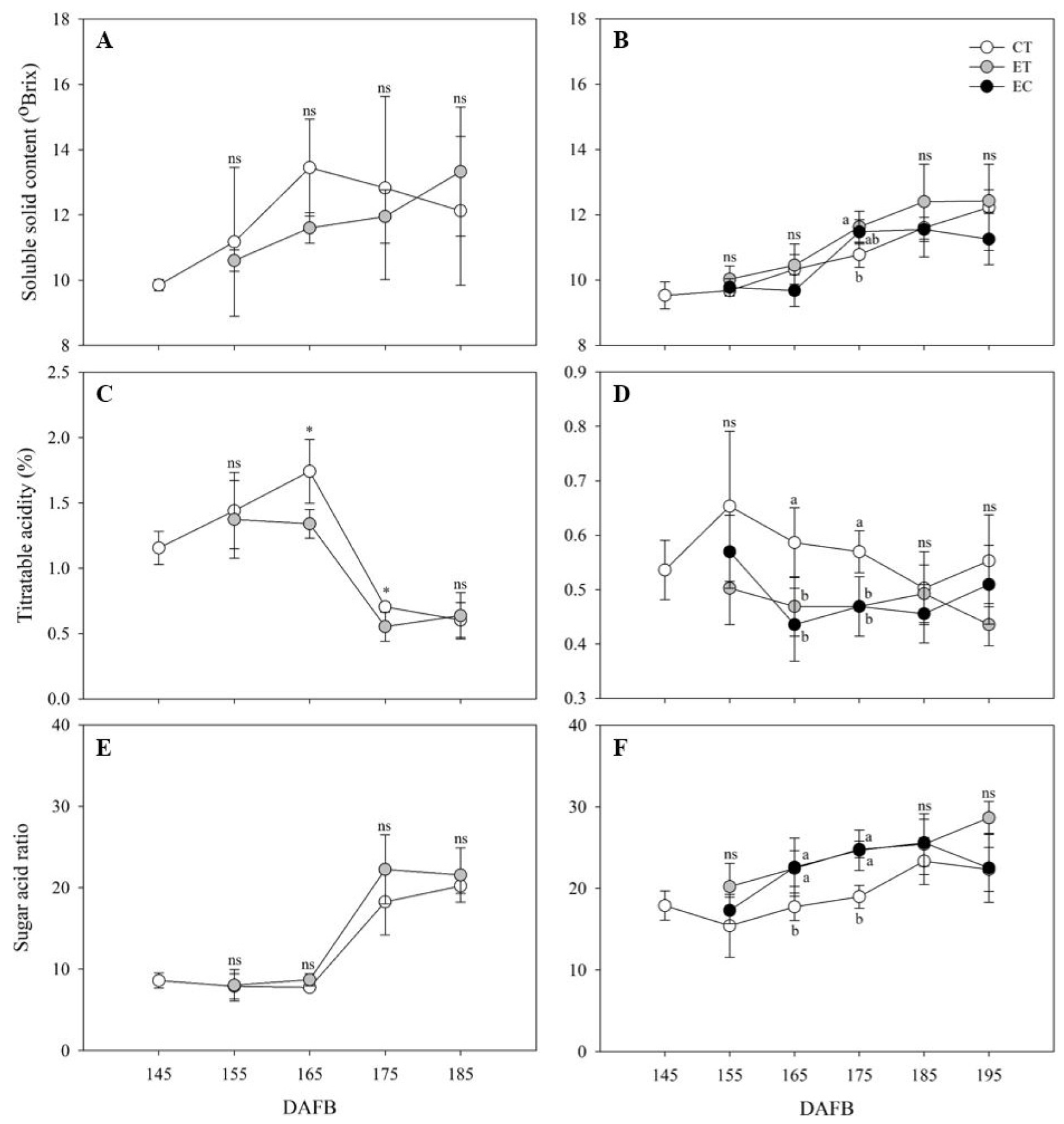
Fig. 6.
Changes of soluble solid content (A and B), titratable acidity (C and D), and sugar acid ratio (E and F) in control (CT), ethephon treated (ET), and ethephon and CaCl2 treated (EC) ‘Fuji’/M.26 fruits collected at different ripening stages in 2020 (A, C, and E) and 2021 (B, D, and F). DAFB, days after full bloom. The bars indicate ± SD (n = 4). Significant differences investigated by t-test are represent to * (p < 0.05), ** (p < 0.01), *** (p < 0.001), and ns (not significance). Different letters indicated significant differences between the treatments at p < 0.05.
고 찰
Ethephon은 ethylene 발생을 촉진하여 식물의 성숙과 노화를 촉진하는 식물 생장조절제로 다양한 원예작물의 재배와 수확 후 관리에 이용되고 있다(Ban et al., 2007). 과수작물의 수확 전 ethephon 처리는 수확시기를 촉진하여 과피 착색을 증진할 수 있으나(Kim and Chung, 2000; Smith and Whiting, 2010), 세포벽 연화를 촉진하여 과실 경도가 감소하는 문제가 발생한다(Dhillon and Mahajan, 2011; Hayama et al., 2006). 본 연구에서는 ‘Fuji’/M.26의 안정적인 수확기 조절 방법을 조사하고자 과피 착색 증진과 과실 성숙 촉진을 위한 ethephon 처리 효과와 ethephon 처리에 따른 경도 감소를 완화하기 위한 ethephon 및 염화칼슘 혼합처리의 효과를 비교하였다.
사과 ‘Fuji’/M.26의 과실 비대생장이 종료된 만개 후 145일에 100 ㎎/L ethephon을 처리한 결과, 처리 10일 후부터 전분함량이 감소하였다(Fig. 1). 이는 ethephon 처리에 의해 ‘Fuji’/M.26의 과실 성숙이 촉진된 것으로 판단된다. 대조구에서 장기저장에 적합한 전분함량을 나타낸 2020년 만개 후 175일과 2021년의 만개 후 185일은 각 해의 적정 수확기로 판단되었다. 2020년과 2021년 적정 수확기의 대조구의 전분함량은 수확 10일 전 ethephon 처리구의 전분함량과 유사하여 100 ㎎/L ethephon 처리가 수확기 조절에 효과적일 것으로 생각되었다.
Ethephon 및 염화칼슘 혼합처리 또한 처리 10일 후부터 ‘Fuji’/M.26의 과실 성숙을 촉진하였다(Fig. 1B). 하지만 전분함량 감소는 ethephon 단용처리구가 ethephon 및 염화칼슘 혼합처리구보다 빨랐다. 세포 내 칼슘 함량이 높으면 원예산물의 호흡과 ethylene 발생을 억제하여 저장성이 향상되는 것으로 알려져 있다(Chung et al., 1993). 본 연구에서 ethephon과 염화칼슘 혼합처리구는 ethephon 단용처리구보다 ethylene 발생량이 적어 상대적으로 성숙이 지연된 것으로 생각된다. 하지만 ethephon 및 염화칼슘 혼합처리구의 만개 후 175일의 전분함량은 만개 후 185일의 대조구와 유사하여(Fig. 1B) ethephon 단용처리 또는 ethephon과 염화칼슘 혼합처리를 통해 사과 ‘Fuji’/M.26의 과실 수확을 10일 정도 촉진할 수 있을 것으로 생각되었다.
사과 ‘Fuji’/M.26의 과피 착색은 ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리에 의해 촉진되었다(Fig. 2). 수확 4주 전 ‘Fuji’ 사과에 100 ㎎/L ethephon 처리는 안토시아닌 생합성을 촉진하여 과피 착색을 증진하는 것으로 보고되었다(Li et al., 2002). 본 연구에서는 수확적기 30~40일 전 100 ㎎/L ethephon 처리를 통해 과피 착색을 촉진할 수 있었다. 2020년 ethephon 처리구는 만개 후 165일에 대조구의 수확기(만개 후 175일) 수준으로 과피 착색이 촉진되었다. 또한, 2021년 ethephon 단용처리구는 ethephon 및 염화칼슘 혼합처리구보다 과피 착색률이 높았으며 두 처리구의 hue 값은 유의적인 차이가 없었다(Fig. 3B). 2021년 만개 후 175일에 ethephon 및 염화칼슘 혼합처리구의 hue 값이 2021년 ‘Fuji’/M.26의 수확기인 만개 후 185일에 대조구의 hue 값과 유사하고 만개 후 175일에 ethephon 단용처리구의 hue 값은 만개 후 185일의 대조구보다 낮아 ethephon 단용처리 또는 ethephon 및 염화칼슘 혼합처리로 과피 착색이 증진된 과실을 10일 정도 조기수확 할 수 있을 것으로 판단되었다.
수확시기별 사과 ‘Fuji’/M.26의 과중은 2021년 수확기인 만개 후 185일을 제외하고 세 처리구 간의 유의적인 차이는 없었다(Fig. 4). 2021년 만개 후 185일에는 ethephon 및 염화칼슘 혼합처리구의 과중이 ethephon 단용처리구의 과중보다 유의적으로 높았다. 만개 후 185일에 ethephon 단용처리구의 과중이 감소 후 195일에 증가한 것은 만개 후 175, 185, 195일 과중의 유의적 차이가 없어 시료수집에 따른 오차로 생각된다.
과실 경도는 2020년 만개 후 175일과 185일에 대조구에서 유의적으로 높게 조사되었으나 2021년에는 세 처리구 간의 유의적인 차이는 없었다. 하지만 대조구와 ethephon 및 염화칼슘 혼합처리구의 경도는 전 시기 동안 0.95 ㎏Ø5 ㎟ 이상의 경도를 유지하였다. 또한, ethephon 및 염화칼슘 혼합처리구는 만개 후 175일부터 가장 높은 평균 경도를 나타내었다(Fig. 5). 이는 염화칼슘 처리에 의해 ethephon에 의한 과실 연화 작용이 억제되고 세포벽 구조가 유지 또는 강화된 것으로 생각된다(Singh et al., 2012). 또한, ethephon 단용처리구는 내생 ethylene 생합성이 촉진되어 과실성숙과 관련된 효소들의 활성화로 세포벽이 연화되어 경도가 감소한 것으로 생각된다(Dhillon and Mahajan, 2011).
당도는 2021년 만개 후 175일을 제외하고 2020년과 2021년 모두 처리구 간의 유의적인 차이는 없었다. 2020년 과실 당도의 표준편차가 큰 이유는 무작위 시료 수집에 따른 과실 간 편차로 생각된다. 2021년 전 수확시기 동안 ethephon 단용처리구는 가장 높은 평균 당도를 나타내었다. Ethephon 단용처리구의 전분함량이 대조구와 ethephon 및 염화칼슘 혼합처리구 보다 빠르게 감소하는 것으로 보아(Fig. 1) ethephon 처리에 의한 전분함량 감소로 가용성 당함량이 증가한 것으로 생각된다. 2021년 만개 후 175일에는 ethephon 단용처리구의 당도는 대조구보다 유의적으로 높았으며, ethephon 및 염화칼슘 혼합처리구는 대조구 및 ethephon 단용처리구와 유의적인 차이는 없었으나 만개 후 185일에 대조구의 당도와 유사하여 전분함량 감소 경향과 일치하였다(Figs. 1, 6B). 칼슘은 과실 발달과 성숙에 중요한 역할을 하며, 수확 전 칼슘처리는 과실성숙을 지연시키고 과실 품질을 유지할 수 있다고 보고되었다(Madani et al., 2016; Zhang et al., 2019). 따라서 ethephon 및 염화칼슘 혼합처리는 ethephon 단용처리보다 과실성숙 촉진효과가 낮은 것으로 생각되었다.
2020년 ‘Fuji’/M.26의 수확기인 만개 후 175일에 ethephon 처리구의 적정산도는 대조구보다 유의적으로 낮아(p < 0.05) 산함량이 감소하며 과실성숙이 촉진된 것으로 생각되었다. 2021년 수확기인 만개 후 185일에 세 처리구의 적정산도는 유의적인 차이가 없었다(Fig. 6C, D). 하지만 2021년 만개 후 175일의 ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구의 적정산도는 대조구보다 유의적으로 낮아 대조구보다 당산비가 높았으며, 수확기 당산비와 유사하였다(Fig. 6F). 따라서 과실 비대생장 종료 후 100 ㎎/L ethephon 단용 처리 또는 100 ㎎/L ethephon 및 0.5% 염화칼슘 혼합처리를 통해 ‘Fuji’/M.26의 과실성숙을 촉진할 수 있음을 확인하였다.
‘Fuji’ 사과는 생육기에 고온이 유지되는 기간이 길고 착색기에 수광이 불량하거나 저온이 늦게 오는 경우 과피 착색이 지연되어 늦게는 12월 초까지 수확한다. 하지만 이와 같이 ‘Fuji’ 사과의 수확이 늦어지면 과실에 밀병현상이 발생하거나 겨울철 동결피해를 받을 수 있어 수확 후 장기저장에 불리하다(Sagong et al., 2013). 또한, 사과나무 재배에 있어 과실 비대생장이 종료되는 만개 후 145일을 전후하여 과피 착색 증진을 위한 도장지 제거와 반사필름 멀칭, 적엽, 착색제 살포작업이 수행된다(Park et al., 2011). 이러한 재배관리는 상품성이 높은 과실 수확을 위해 수행되지만 과도한 노동력과 경영비 투입을 야기한다.
본 연구를 통해 과실 비대생장 종료 후 100 ㎎/L ethephon 단용처리 또는 100 ㎎/L ethephon 및 0.5% 염화칼슘 혼합처리로 ‘Fuji’/M.26 사과의 착색을 증진하고 과실성숙을 수확적기보다 10일 촉진할 수 있었다. 하지만 ethephon 단용처리는 과실 경도가 낮아 식미가 불량해지는 단점이 있어 100 ㎎/L ethephon 및 0.5% 염화칼슘 혼합처리를 통해 상품성이 높은 과실을 수확적기보다 10일 조기 수확할 수 있을 것으로 생각된다. 또한, 이와 같은 수확기 조절 방법을 활용하여 수확 지연으로 수체 내 저장양분의 과다 소비로 인한 다음해 해거리 증상과 봄철 냉해 및 동해를 예방할 수 있을 것으로 생각된다.
적 요
만생종 ‘Fuji’ 사과(Malus × domestica)는 과피 착색에 따라 수확시기가 가변적인 품종이다. Ethephon은 ethylene 발생을 촉진하는 식물생장조절제로 사과나무를 포함한 호흡급등형 과수작물의 과실 성숙과 관련된 생리적 반응을 유도한다. 본 연구는 ‘Fuji’/M.26 사과의 경제적이고 안정적인 수확기 조절 방법을 제시하고자 과실 비대생장 종료 후 ethephon 단용처리와 ethephon 및 염화칼슘 혼합처리가 과실특성에 미치는 영향을 조사하였다. 과실 비대생장이 종료된 만개 후 145일에 100 ㎎/L ethephon과 100 ㎎/L ethephon 및 0.5% 염화칼슘 혼합용액을 분무처리 후 10일 주기로 과실특성을 조사하였다. Ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구 모두 만개 후 155일부터 전분함량이 감소되어 과실성숙이 촉진되었으며, 수확 10일 전 전분함량은 대조구의 수확기 전분함량과 유사하였다. 과피 착색은 ethephon 및 염화칼슘 혼합처리구에서 ethephon 단용처리구보다 착색률이 낮았으나 대조구보다는 높았다. 과실 평균 경도는 ethephon 단용처리구가 가장 낮았으며, ethephon 및 염화칼슘 혼합처리구는 대조구와 유사하였다. 수확기 당산비는 세 처리구 간에 유의한 차이는 없었으나 수확 10일 전 ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구의 적정산도가 대조구보다 유의적으로 낮았다. 특히 2021년 수확 10일 전 ethephon 단용처리구와 ethephon 및 염화칼슘 혼합처리구의 당산비는 수확기 당산비와 유사하였다. 따라서 ‘Fuji’/M.26의 과실 비대생장이 종료된 시점에 100 ㎎/L ethephon과 0.5% 염화칼슘 혼합처리를 통해 과실성숙과 과피 착색을 수확기보다 10일 촉진할 수 있을 것으로 생각된다.
