

서 언
며느리밥풀속(Melampyrum L.)은 꿀풀목(Lamiales), 현삼과(Scrophulariaceae)에 속하는 1년생 초본식물로(Beauverd, 1916) 반기생하는 생태학적 특성과 분자생물학적 연구에 의해 현삼과가 아닌 열당과(Orobanchaceae)에 포함되기도 한다(Bae, 2011). 며느리밥풀속 식물들은 북반구의 온대지역에 약 40여종이 분포하는 것으로 알려져 있으며(Shin, 1992), 대다수 종이 유럽지역에 집중분포하고, 북미지역과 동아시아지역에는 일부 종이 분포한다(Kitamura, 1941; Nakai, 1908, 1917; Park, 1974; Soo, 1926, 1927; Tuyama, 1941). 최근 Lee et al. (2016)은 강원도를 중심으로 며느리밥풀속의 서식지 특성 및 개미와의 공생관계에 관한 연구를 진행한 바 있으며, 본 속에 속하는 종들의 대부분은 건조한 수풀, 초원과 목초지에 주로 생육하고, 일부 양지바른 산기슭과 능선, 고산지대에 분포하는 반면, 습기가 많은 수풀과 음지, 늪에는 거의 생육하지 않는 것으로 보고되었다(Kitamura, 1941; Nakai, 1917; Soo, 1926, 1927; Tuyama, 1941; Yeom, 2009).
한국산 며느리밥풀속에 관한 연구는 Nakai (1907)가 처음 1종 2변종(Melampyrum roseum Maxim., M. roseum var. japonicum Franch. & Sav., M. roseum var. setaceum Maxim.)이 한반도에 분포한다고 보고하였으며, 이후 잎의 형태, 포의 색, 포의 가장자리 강모 형태, 분지의 형태와 습성 등을 종합하여 5종 3변종 1품종으로 정리하였다(Nakai, 1917). 이후 한반도에 분포하는 며느리밥풀속은 이명과 변종의 처리에 따라 분류군의 수도 달리 보고되어 왔다. Chung et al. (1937)은 한반도에 분포하는 며느리밥풀속을 국명과 함께 5종으로, Park (1949)은 7종 6변종 4품종으로 보고하였으며, Chung et al. (1949)은 5종 2변종으로, Park (1974)은 4종 2변종으로 보고하였다. Lee and Yamazaki (1983)는 한국의 현삼과 식물의 학명을 정리하면서 그동안 정명으로 사용되어왔던 M. setaceum var. latifolium Nakai을 M. setaceum의 이명으로, M. ciliare Miquel을 M. roseum var. japonicum의 이명으로 처리하고, M. roseum var. ovalifolium Nakai ex Beauverd과 M. setaceum var. nakaianum (Tuyama) T. Yamaz.을 추가하여, 한국에 분포하는 며느리밥풀속을 M. roseum을 포함한 2종 3변종 1품종(M. roseum var. ovalifolium f. albiflorum Nakai ex T. Lee)으로 보고하였다. Lee (1996)는 며느리밥풀속의 학명들을 재정리하면서 Lee and Yamazaki (1983)의 보고에 M. roseum var. hirsutum Beauv.와 M. roseum var. japonicum f. leucanthum Nakai을 추가하여 2종 4변종 2품종으로 보고하였으며, Lee (2006)는 흰애기며느리밥풀(M. setaceum f. albiflorum Y. Lee)을 추가하여 2종 4변종 3품종의 한국산 며느리밥풀속을 기재하였다. Kim and Yun (2012)은 신종으로 발표한 긴꽃며느리밥풀(Melampyrum koreanum K. J. Kim & S. M. Yun)을 더한 3종 4변종 3품종으로 보고하였다. 이에 본 연구는 한국에 분포한다고 알려진 며느리밥풀속 3종 4변종 3품종 중에서 분류군의 명확한 실체와 분포가 확인되지 않은 털며느리밥풀, 흰수염며느리밥풀, 흰알며느리밥풀, 흰애기며느리밥풀 등을 제외한 3종 3변종(꽃며느리밥풀; M. roseum, 수염며느리밥풀; M. roseum var. japonicum, 알며느리밥풀; M. roseum var. ovalifolium, 애기며느리밥풀; M. setaceum, 새며느리밥풀; M. setaceum var. nakaianum, 긴꽃며느리밥풀; M. koreanum)을 대상으로 본 연구를 수행하였으며, Moon and Jang (2020)의 조사방법을 참고하여 기술하였다.
그동안 한국산 며느리밥풀속의 분류형질에 관하여 Lee (1996)와 Lee (2003), Nakai (1917), Park (1974) 등의 학자들이 제시한 잎의 형태와 털의 유무, 화서의 길이, 포 가장자리의 강모 수, 포 간 간격, 개화기 포의 색, 꽃받침의 털 유무 등을 검색형질로 분류군의 동정이 이루어져 왔으나, 며느리밥풀속의 반기생성 생육 특성에 따라 지역 간 변이로 인해 보다 명확한 검색형질의 설정이 필요한 실정이다. 이에 기존에 알려진 검색 형질을 포함한 주요 외부형태학적 형질을 조사하고, 이를 통해 한국산 며느리밥풀속 6분류군의 동정에 유용한 검색 형질들을 재검토하고자 하였다.
재료 및 방법
본 연구에 사용된 재료는 2012년 6월부터 2021년 10월까지 개화기와 결실기를 중심으로 채집한 건조표본과 성숙한 개체를 FAA에 고정한 액침표본, 그리고 국립수목원 식물표본실(KH), 안동대학교 표본실(ANH), 국립백두대간수목원 표본관(KBA)에 소장되어있는 표본 등을 대상으로 하여 각 분류군별로 30개체 이상 총 180개체 이상을 조사하였다. 정량적 및 정성적 형질은 육안 또는 해부현미경(Olympus AX-70, Japan; Olympus DP2-BSW, Germany)을 사용하여 관찰 및 촬영하였다. 연구에 사용된 표본정보는 Appendix 1에 기재하였다.
결 과
한국산 며느리밥풀속(Melampyrum L.) 6분류군의 주요 외부형태학적 형질을 관찰하고 기재하였다. 잎의 형태, 포의 형태, 포 가장자리 강모의 분포 위치와 수는 유용한 검색형질로 확인되었으며, 줄기의 횡단면 형태, 잎의 크기, 포의 기부 형태, 화관의 색과 길이, 열매의 형태, 종자의 크기 등 질적, 양적 형질기재는 Table 1, 2에 제시하였다.
Table 1.
Qualitative characters in Korean Melampyrum
줄기
줄기는 6분류군 모두 직립하였으며, 분지의 수는 생육 특성에 따라 1-9개로 관찰되었다. 횡단면의 형태와 털의 분포에 따라 분류군들이 구분되었다(Table 2).
Table 2.
Quantitative characters in Korean Melampyrum (min (mean) max)
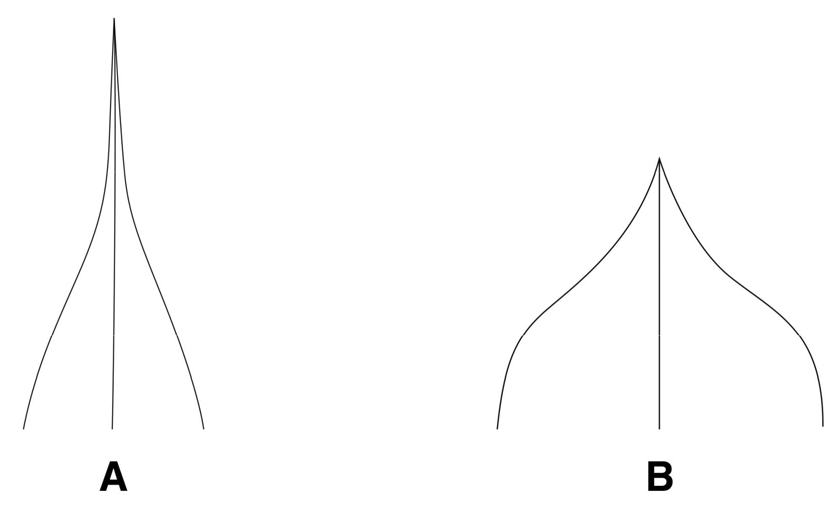
줄기 횡단면의 형태
(1) 사각형(sub-quadrangular): 꽃며느리밥풀, 알며느리밥풀, 수염며느리밥풀, 긴꽃며느리밥풀의 4분류군에서 둔각을 가진 사각형의 형태로 관찰되었다(Fig. 1A).
(2) 원형(circular): 애기며느리밥풀과 새며느리밥풀의 2분류군에서 관찰되었다(Fig. 1B).

줄기의 털 분포
꽃며느리밥풀, 알며느리밥풀, 수염며느리밥풀, 긴꽃며느리밥풀이 능선을 따라 분포하였으며, 새며느리밥풀, 애기며느리밥풀은 줄기 전체에 걸쳐 털이 분포하는 것으로 관찰되었다(Fig. 2).

잎
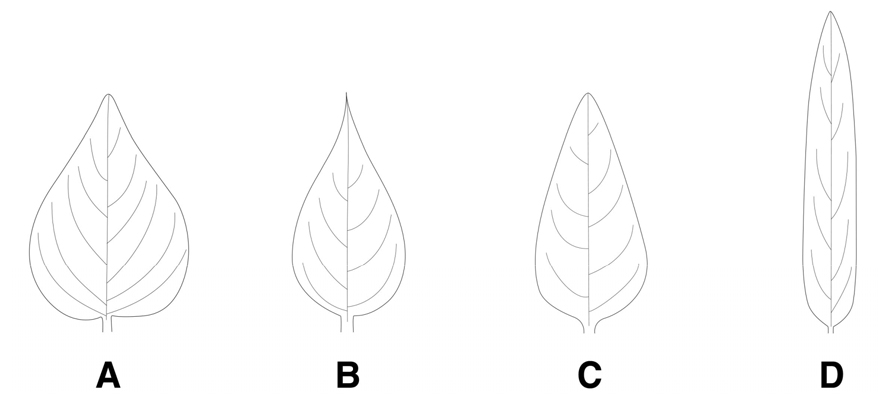
잎의 형태
(1) 난형(ovate): 잎의 선단부에서 기부로 갈수록 상대적으로 폭이 넓어지는 달걀모양의 형태로 수염며느리밥풀에서 관찰되었다(Fig. 3A).
(2) 난상피침형(ovate-lanceolate): 기부는 달걀모양으로 잎의 선단부로 갈수록 뾰족하게 좁아지는 형태이며, 꽃며느리밥풀과 알며느리밥풀에서 관찰되었다(Fig. 3B).
(3) 피침형(lanceolate): 기부에서부터 1/3정도 되는 부분의 폭이 가장 넓고 선단부으로 갈수록 창처럼 좁아지는 형태로 새며느리밥풀과 긴꽃며느리밥풀에서 관찰되었다(Fig. 3C).
(4) 선형(linear): 잎의 길이가 폭보다 2배 이상 길고, 잎 가장자리가 평행하면서 좁아지는 형태로 애기며느리밥풀에서 관찰되었다(Fig. 3D).
잎의 크기
잎의 너비는 수염며느리밥풀이 평균 15.5 ㎜와 20.8 ㎜로 가장 넓게 나타났고, 애기며느리밥풀이 평균 3.5 ㎜와 4.5 ㎜로 가장 좁게 나타났으며, 잎의 길이는 새며느리밥풀이 41.4 ㎜와 59.1 ㎜로 가장 길게 나타났다(Table 2).
잎의 털
잎에서의 털의 유무와 분포는 모든 분류군이 맥을 포함한 잎의 표면과 이면 전체에 걸쳐 털이 분포하고 있으며, 수염며느리밥풀은 타 분류군과 확연히 구별되는 다세포의 긴 백색 연모가 관찰되었다.
화서
화서의 길이는 15.3-260.6 ㎜의 범위로 측정되었다. 긴꽃며느리밥풀이 평균 152.3 ㎜, 꽃며느리밥풀이 평균 107.8 ㎜, 알며느리밥풀이 75.5 ㎜로 측정되었으며, 애기며느리밥풀과 새며느리밥풀의 화서 길이는 평균 32.5 ㎜와 40.1 ㎜의 짧은 형태로 측정되었다(Table 2). 특히 수염며느리밥풀은 개화기에도 포가 화서를 감싸듯 뭉쳐져 있는 형태로 관찰되었다.
포
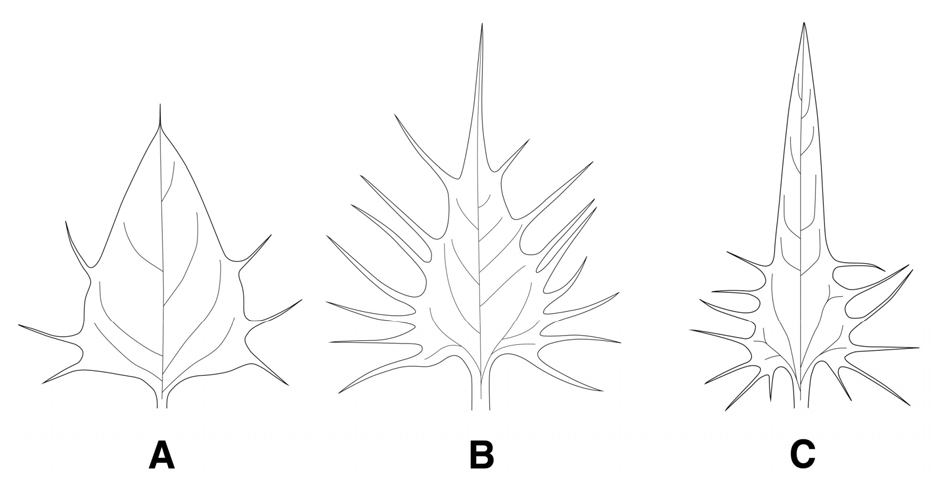
포의 형태
(1) 넓은삼각형(wide-triangle): 꽃며느리밥풀, 수염며느리밥풀, 긴꽃며느리밥풀에서 관찰되었다(Fig. 4A).
(2) 삼각형(triangle): 알며느리밥풀과 새며느리밥풀에서 관찰되었다(Fig. 4B).
(3) 좁은삼각형(narrow-triangle): 애기며느리밥풀에서만 관찰되었다(Fig. 4C).
포의 기부 형태
(1) 평저(truncate): 포의 기부가 수평으로 평평한 모양으로 꽃며느리밥풀, 새며느리밥풀, 긴꽃며느리밥풀에서 관찰되었다(Table 1).
(2) 심장저(cordate): 포의 기부가 심장의 밑부분 모양으로 알며느리밥풀, 수염며느리밥풀, 애기며느리밥풀에서 관찰되었다(Table 1).
포의 강모
포의 가장자리에 있는 강모의 분포 위치(포의 중·하부)와 수는 꽃며느리밥풀이 1-6쌍, 긴꽃며느리밥풀은 0-4쌍의 강모가 분포하는 반면, 알며느리밥풀은 포 전체에 걸쳐 5-10쌍, 애기며느리밥풀 4-8쌍, 새며느리밥풀이 4-7쌍의 강모 수로 관찰되었다. 수염며느리밥풀은 포 전체에 걸쳐 가장 많은 7-13쌍의 강모 수가 관찰되었다(Table 2).
포의 간격
긴꽃며느리밥풀과 꽃며느리밥풀의 포 간격은 평균 8.0 ㎜ 이상이며, 애기며느리밥풀은 평균 3.5 ㎜로 가장 좁은 간격으로 관찰되었다(Table 2).
포의 색
개화기 포의 색을 조사한 결과, 애기며느리밥풀과 새며느리밥풀은 포의 색이 녹색에서 자홍색으로 변하는 것으로 관찰되었으며, 새며느리밥풀은 애기며느리밥풀과 달리 종자 결실 후 포의 색이 연한 홍색으로 변색되었다. 꽃며느리밥풀, 알며느리밥풀, 수염며느리밥풀, 긴꽃며느리밥풀은 개화기에도 포의 색이 대부분 녹색을 유지하였으나, 일부 조사지역에 분포하는 꽃며느리밥풀과 수염며느리밥풀에서 포의 색이 자홍색을 띠는 변이개체가 관찰되었다(Fig. 5).

꽃
화관의 길이와 색
하부 화판의 정단부까지 측정한 화관의 길이는 긴꽃며느리밥풀이 평균 37.6 ㎜의 화관 길이로 타 분류군의 평균 15.4- 17.9 ㎜와 확연히 구분되었으며, 화관 색에서도 긴꽃며느리밥풀은 연한 자주색으로 관찰되어 타 분류군들의 짙은 자홍색과 구별되었다(Table 1-2).
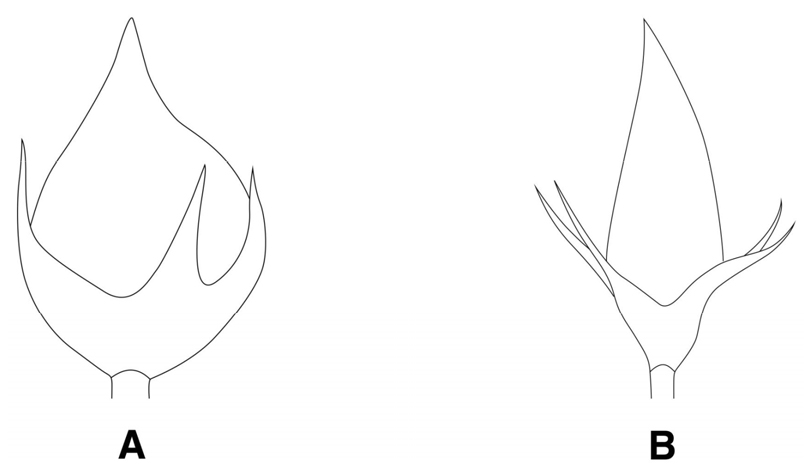
꽃받침 열편과 정단부 돌기의 형태
꽃며느리밥풀은 까락형의 예두로 나타나 엽침형의 점첨두를 보인 타분류군과 구별되었다(Fig. 6).
꽃받침의 털
모든 분류군에서 맥을 포함한 꽃받침 전체에 털이 분포하는 것으로 확인되었으며, 수염며느리밥풀은 타 분류군과 구별되는 다세포의 긴 백색 연모가 관찰되었다.
열매
열매는 삭과이며, 모든 분류군이 포배열개 하였다.
열매의 형태
(1) 난형(ovate): 꽃며느리밥풀, 애기며느리밥풀, 수염며느리밥풀, 알며느리밥풀과 긴꽃며느리밥풀은 난형으로 관찰되었으며, 특히 긴꽃며느리밥풀의 너비와 길이는 평균 4.8 ㎜, 9.5 ㎜로 분류군 중 가장 큰 것으로 관찰되었다(Fig. 7A, Table 2).
(2) 피침형(lanceolate): 열매의 형태에서 새며느리밥풀에서만 피침형으로 관찰되었다(Fig. 7B, Table 1).
종자
종자의 형태
6분류군 모두 가종피를 가지고 있는 장타원형으로 나타났으며, 종자의 색은 밝은 황색으로 관찰되었다(Fig. 8).

종자의 너비와 길이
종자의 너비는 애기며느리밥풀이 평균 1.4 ㎜로 가장 낮게 나타났고, 길이는 알며느리밥풀과 수염며느리밥풀이 평균 3.6 ㎜로 낮게 측정되었으며, 긴꽃며느리밥풀이 평균 1.9 ㎜와 5.2 ㎜로 타 분류군 보다 높게 나타났다(Table 2).
열매 당 결실된 종자 수
새며느리밥풀이 평균 1.35개로 가장 높은 결실 종자 수를 나타낸 반면, 긴꽃며느리밥풀은 가장 낮은 0.96개로 나타났다(Table 2).
이상과 같은 외부형태학적 분류형질을 바탕으로 한국산 며느리밥풀속 6분류군의 검색표를 작성하였다.
한국산 며느리밥풀속(Melampyrum L.)의 검색표
1.화관의 길이가 3 ㎝ 이상이다.....................................................................................................긴꽃며느리밥풀(M. koreanum)
1.화관의 길이가 3 ㎝ 이하이다.
2.잎은 난형 또는 난상피침형이다.
3.줄기, 잎, 포, 꽃받침에 긴 백색 연모가 있다........................................................수염며느리밥풀(M. roseum var. japonicum)
3.줄기, 잎, 포, 꽃받침에 긴 백색 연모가 없다.
4.포는 넓은삼각형이며, 기부는 평저이고, 가장자리 중·하부에 걸쳐 1-6쌍의 강모가 있다..........................................................
.........................................................................................................................................꽃며느리밥풀(M. roseum var. roseum)
4.포는 삼각형이며, 기부는 심장저이고, 가장자리 전체에 걸쳐 5-10쌍의 긴 강모가 있다............................................................
...................................................................................................................................알며느리밥풀(M. roseum var. ovalifolium)
2.잎은 선형 또는 피침형이다.
5.잎은 선형이고, 포는 좁은삼각형이며, 기부는 심장저, 열매의 형태는 난형이다........................................................................
..............................................................................................................................애기며느리밥풀(M. setaceum var. setaceum)
5.잎은 피침형이고, 포는 삼각형이며, 기부는 평저, 열매의 형태는 피침형이다...........................................................................
...............................................................................................................................새며느리밥풀(M. setaceum var. nakaianum)
고 찰
한국산 며느리밥풀속 6분류군의 분류학적 실체를 파악하기 위하여 주요 외부형태학적 형질를 파악하였다. 조사 결과, 잎의 형태, 포의 형태, 포 가장자리 강모의 분포 위치와 수는 유용한 검색형질로 확인되었으며, 줄기의 횡단면 형태와 털의 분포 양상, 포의 기부 형태, 꽃받침 열편과 정단부 돌기의 형태, 화관의 색과 길이는 새로운 검색형질로 나타났다(Table 1, 2). 그러나 기존의 검색형질로 알려진 분지의 형태, 잎에서의 털의 유무, 화서의 길이, 포 간 간격, 꽃받침에서의 털의 유무, 개화기 포의 색은 폭넓은 변이로 인해 검색형질로는 부적합한 것으로 나타났다.
잎의 형태는 그동안 한국산 며느리밥풀속의 주요 검색형질로 이용되어왔으나 학자들에 따라 분류군이 서로 달리 기재되어 많은 혼선을 가져온 형질이었다. 잎의 형태는 지금까지 난형으로 알려진 알며느리밥풀은 꽃며느리밥풀과 함께 난상피침형(ovate-lanceolate)으로, 피침형과 난상피침형으로 알려진 수염며느리밥풀은 난형(ovate)으로 나타났으며, 새며느리밥풀과 긴꽃며느리밥풀은 피침형(lanceolate), 애기며느리밥풀은 타 분류군과 확연히 구별되는 선형(linear)으로 확인되었다(Fig. 3, Table 1). 또한, 포의 형태도 난형 또는 삼각상피침형으로 알려져 왔으나 넓은삼각형(wide-triangle), 삼각형(triangle), 좁은삼각형(narrow-triangle)의 3가지 형태로 관찰되었으며(Fig. 4, Table 1), 포의 가장자리 강모의 분포 위치와 수에서 꽃며느리밥풀과 긴꽃며느리밥풀은 포의 중·하부에 평균 5쌍 미만의 강모가 분포하는 것으로 조사되어, 포의 전체에 걸쳐 평균 5-10쌍의 강모가 분포하는 타 분류군과 구별되는 유용한 검색 형질로 확인되었다(Table 2).
줄기의 분지 형태와 수는 본 조사 결과에서 애기며느리밥풀이 타 분류군들의 평균 3-4개 보다 많은 평균 6개의 분지를 형성하였으나 Nakai(1917)가 제시한 검색형질 중 분지의 수는 생육상태에 따라 동일한 분류군 내에서도 개체 간 큰 변이들이 관찰되었으므로 검색 형질로는 부적합한 것으로 판단되었다. 잎에서의 털의 유무는 모든 분류군에서 맥을 포함한 잎의 표면과 이면 전체에 털이 분포하는 것으로 확인되었으며, 꽃받침에서의 털도 모든 분류군에서 관찰되어 검색 형질로는 부적합한 것으로 나타났다. 다만 수염며느리밥풀은 타 분류군들과 확연히 구별되는 긴 백색 연모가 잎과 꽃받침에 밀생하는 것으로 관찰되어 수염며느리밥풀과 타 분류군을 구별하는 유용한 검색 형질로 확인되었다. 화서의 길이와 포 간 간격은 꽃며느리밥풀과 알며느리밥풀 두 분류군을 구별하는 검색 형질로, 꽃며느리밥풀이 알며느리밥풀에 비해 긴 화서와 넓은 포 간 간격을 가지는 것으로 알려져 있었으나, 조사 결과, 화서의 길이는 동일분류군에서도 변이차이가 심하게 나타났고, 포 간 간격도 화서의 길이에 따라 영향을 받는 것으로 관찰되어 검색 형질로는 부적합한 것으로 확인되었다(Table 2). 개화기 포의 색은 Nakai (1917)가 한국산 며느리밥풀속의 검색 형질로 처음 제시한 이후 가장 유용하고 명확한 검색 형질로 알려져 왔으며, 개화기 포의 색에 따라 녹색을 띠는 꽃며느리밥풀, 알며느리밥풀, 수염며느리밥풀 등의 M. roseum계열과 자홍색을 띠는 애기며느리밥풀과 새며느리밥풀의 M. setaceun계열로 크게 구분되어왔다. 그러나 본 조사 결과, M. roseum계열에 속한 꽃며느리밥풀과 수염며느리밥풀의 일부 조사지에서 녹색이 아닌 자홍색을 띠는 포를 가진 변이개체들이 다수 관찰되어 개화기 포의 색은 검색 형질로 부적합한 것으로 나타났다(Table 1).
줄기의 횡단면을 조사한 결과, M. setaceun계열인 애기며느리밥풀과 새며느리밥풀은 원형의 형태로 관찰되어 둔각을 가진 사각형의 형태로 확인된 M. roseum계열과 구별되었으며(Fig. 1), 줄기에서의 털의 분포 양상도 애기며느리밥풀과 새며느리밥풀은 줄기 전체에 걸쳐 털이 분포하고 있는 것으로 확인되어 줄기의 능선을 따라 털이 분포하고 있는 타 분류군과 구별되는 유용한 검색 형질로 밝혀졌다(Fig. 2). 포의 기부 형태는 본 조사에서 꽃며느리밥풀, 새며느리밥풀, 긴꽃며느리밥풀이 평저로 나타나 알며느리밥풀, 수염며느리밥풀, 애기며느리밥풀의 심장저와 구별하는 새로운 검색형질로 확인되었다(Table 1). 꽃받침 열편의 형태는 Beauverd (1916)와 Yamazaki (1954)에 의해 지역에 따른 변이가 심하여 검색형질로 부적합하다고 보고하였으며, 한국산 며느리밥풀속의 검색형질로는 언급되지 않았던 형질이다. 조사 결과, 꽃받침 열편과 정단부 돌기의 형태에서 꽃며느리밥풀은 까락형의 짧은 정단부 돌기를 가진 예두(acute)로 나타나 타 분류군의 엽침형 정단부 돌기를 가진 점첨두(acuminate)와 구분되는 검색형질로 확인되었다(Table 1). 열매의 형태에서는 새며느리밥풀이 타 분류군의 난형과 구분되는 피침형의 열매 형태를 나타내어 새로운 검색형질로 확인되었다. 화관의 길이는 긴꽃며느리밥풀이 평균 37.6 ㎜로 가장 높게 나타났으며, 타 분류군의 평균 화관 길이인 15.4-17.9 ㎜와 확연히 구분되었다(Table 2).
한편, 흑산도와 홍도에서 채집된 꽃며느리밥풀 개체들은 평균 24.8 ㎜의 화관 길이를 나타내 내륙에서 채집한 평균 16 ㎜와 큰 차이를 나타냈다. 그러나 화관의 길이를 제외한 털의 형태, 잎의 형태, 포의 형태와 가장자리 강모의 위치와 수 등의 형질들이 꽃며느리밥풀의 변이 범위 내에 위치하는 것으로 확인되어 본 연구에서도 Yamazaki (1954)의 연구와 같이 꽃며느리밥풀로 동정하였다. 이러한 동일 분류군 내의 화관 길이의 차이는 자생지의 기후와 지역적 환경에 의한 다양한 변이가 발생할 수 있으며, 향후 지속적인 연구가 필요하다고 판단된다.
적 요
한국산 며느리밥풀속(Melampyrum L.) 6분류군(M. roseum, M. roseum var. japonicum, M. roseum var. ovalifolium, M. setaceum, M. setaceum var. nakaianum, and M. koreanum)을 대상으로 외부형태학적 형질을 관찰하였다. 조사 결과, 잎의 형태, 포의 형태, 포 가장자리 강모의 분포 위치와 수는 유용한 검색형질로 확인되었으며, 줄기의 횡단면 형태와 털의 분포, 포의 기부 형태, 꽃받침 열편과 정단부 돌기의 형태, 화관의 색과 길이는 유용한 검색형질로 나타났다. 이러한 외부형태학적 형질을 바탕으로 한국산 며느리밥풀속의 검색표를 제시하였다.
