

서언
측백나무과(Cupressaceae Bartling)는 약 20속 125분류군이 전세계 냉온대지역에 널리 분포한다. 눈측백속(Thuja L.)은 극동아시아(한국, 중국, 일본)와 북아메리카에 5분류군이 자생한다(Page, 1990; Farjon, 2005). 우리나라에는 눈측백(Thuja koraiensis Nakai), 측백나무(Thuja orientalis L.) 2분류군이 보고되어있다(Lee, 1996).
눈측백은 IUCN (2015)의 적색목록 범주와 기준으로 VU (Vulnerable)등급 취약종으로 분류하고 있으며, 우리나라와 중국에서도 눈측백을 VU등급(Fu and Jin, 1992; Korea National Arboretum, 2008; National Institute of Biological Resources, 2012; Wang and Xie, 2004)으로 구분하고 있다. 또한, 식물구계학적 특정식물 Ⅴ등급(Kim, 2000), 지구온난화에 민감한 지표식물(Kong, 2005) 및 한반도 기후변화 적응 대상식물 300 (Korea National Arboretum, 2010)으로 지정되었다.
최근의 기온 상승은 가뭄, 홍수 등 극한 기상현상의 발생 증가와 빙하면적의 감소로 인한 해수면 상승 등을 야기한다. 이러한 지구 물리적환경 변화는 자연생태계 전반에 광범위한 영향을 끼치고 있다(IPCC, 2007). 식물종은 변화하는 환경에 적응하겠지만 그렇지 못한 식물들은 멸종하게 될 것으로 예상하고 있다(Tellez et al., 2007). 또한, 온대성 식물의 분포는 확산되는 반면 한대성 식물의 분포는 축소되어 저지대 식물이 고산지대로 이동한다는 연구가 보고되고 있다(Kullman, 2002; Meshinev et al., 2000; Moiseev and Shiyatov, 2003).
따라서 지구온난화 영향에 따른 생물종의 분포, 환경요인 등 관계를 밝히는 연구가 중요시 되고 있으며(Woodward, 1987), 멸종위기에 처한 식물을 대상으로 정량적 분석을 통한 종 보전의 필요성이 대두됨에 따라 멸종위기식물에 대한 연구가 이루어지고 있다(Ghazaly, 2007; Golicher et al., 2008; Newton, 2007).
최근 우리나라에서도 왕제비꽃(Song et al., 2010), 줄댕강나무(Kim et al., 2010) 금강제비꽃(Seo and Yoo, 2011), 호랑가시나무(Kwon et al., 2011), 섬잣나무-솔송나무(Choet al., 2011), 너도밤나무(Cheon et al., 2012), 자란(So and Song, 2012), 애기송이풀(Byun et al., 2013), 갯금불초(Kim et al., 2013) 등에 대한 연구가 활발히 진행되고 있다.
눈측백에 대한 선행연구는 I-SSR표지자에 의한 유전변이 연구(Yang et al., 2009), 삽목번식에 관한 연구(Song et al., 2006), 생물 지리적으로 한반도 고산식물 분포(Kong, 2002), 침엽수종 분포(Kong, 2004) 등 분포학적 연구는 진행되었으나 생태학적 접근에 관한 연구는 전무한 실정이다.
따라서 본 연구는 희귀식물인 눈측백에 대한 군집구조 및 생육환경과의 관계를 밝히고 효율적인 보전 및 관리의 생태적 증거자료를 제공하고자 한다.
재료 및 방법
조사지역 및 현지조사
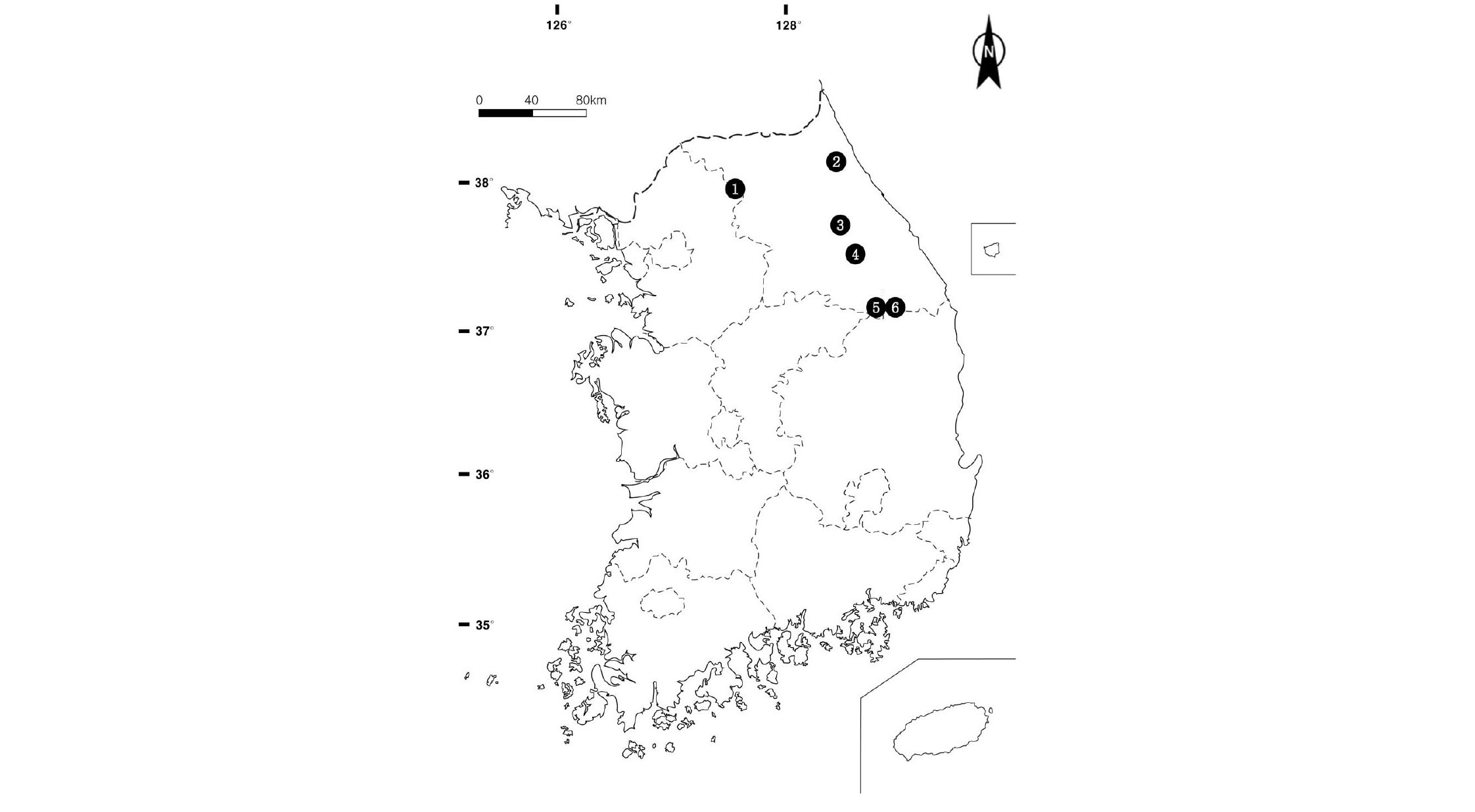
가리왕산(4개소), 계방산(3개소), 장산(3개소), 태백산(2개소), 화악산(2개소), 설악산(8개소)의 눈측백 자생지의 층위별 식생구조 분석을 위해 10 m × 10 m, 15 m × 15 m의 방형구 22개소를 설치하였다(Fig. 1). 현지조사는 2011년 4월부터 2012년 10월까지 눈측백의 생육특성을 감안하여 방형구 내 집중적으로 분포하는 하층 식생 조사구 2개(2 m × 2 m)를 설치하여 개체수와 피도값을 조사하였다.
식생조사는 수관의 위치에 따라 교목․아교목․관목층으로 구분하여 Braun-Blanquet (1965)의 통합우점도를 이용하였으며, 임분구조의 정량적 분석을 위해 1 ㎝이상 개체목의 흉고직경(DBH)을 측정하였다.
식물동정은 Lee (1996, 2006a, 2006b), Lee (1980, 2003a, 2003b), Park (2009), Korean Fern Society (2005), Korea National Arboretum (2011)을 이용하였다. 학명 및 국명은 국가표준식물목록(Korea National Arboretum, 2015)에 준하였다.
자료분석 프로그램은 종간 상관 분석은 SPSS PASW Statistics (ver. 18.0), 유집분석, NMS 분석, 지표종 분석은 McCune and Mefford (2006) PC-ORD (ver. 5.17)를 이용하였다.
유집분석의 경우 교목층과 아교목층의 흉고단면적(Basal area)을 이용하였고, 각 조사구의 거리 척도는 Euclidean (Pythagorean)으로 군집분류는 Ward’s 방법으로 분석하였다.
종간 상관분석을 위하여 각 조사구의 식생조사 자료를 이용하였다. 층위별 우점종 분석 및 종간 상대적 우세를 비교하기 위하여 Curtis and McIntosh (1951)의 중요치(I.V.: Importance Value)를 백분율로 나타낸 상대우점치(Brower and Zar, 1987)를 상층(교목․아교목층)과 하층(관목․초본층)으로 구분하여 분석하였다. 상대우점치(Importance Percentage)는 (상대밀도+상대피도+상대빈도) / 3으로 계산하였으며, 상대밀도는 개체수 자료를 이용하였고, 상대피도는 흉고단면적을 이용하였다.
NMS 분석(다차원척도법: Non-metric Multidimensional Scaling)의 경우 식생조사 자료와 환경조사 자료를 이용하여 2차원 상에 배열하였다. 유집분석으로 분류된 군집에 대해 MRPP검정(다수응답순열절차: Multi-Response Permutation Procedures)을 하였다. MRPP 검정은 집단 분포의 정규성, 동질성 가정을 피할 수 있는 생태학적 자료에 적합한 비모수 분석방법으로 각 집단 간 종조성의 차이를 검증 할 수 있다(McCune and Grace, 2002; McCune and Mefford, 2006).
상층 식생의 흉고단면적과 하층 식생의 조사구별 출현종 통합우점도 계급(피도범위 중앙치로 변환 후 출현종 상대값 합을 100으로 환산)을 분석에 이용하였다.
지표종 분석(indicator species analysis)은 상층 및 하층 식생으로 구분하여 군집에서 유의성이 인정되는 종을 분석하였으며, 환경요인은 고도, 경사, 노암율, 토양 유기물, 낙엽층 깊이와 흉고단면적 등을 이용하였다.
결과 및 고찰
관속식물상 및 환경요인
22개의 방형구 내에서 조사된 관속식물상은 총 51과 101속 136종 1아종 11변종 2품종으로 총 150분류군이 분포하는 것으로 조사되었다. 양치식물은 6과 10속 19분류군, 나자식물은 3과 5속 6분류군, 피자식물 중 쌍자엽식물은 37과 70속 99분류군, 단자엽식물은 5과 16속 26분류군이다. 설악산지역 방형구 내에서 87분류군으로 종풍부도가 가장 높은 것으로 조사되었으며, 장산 51분류군, 가리왕산 41분류군, 계방산 40분류군, 태백산 31분류군, 화악산 21분류군 순으로 조사되었다.
방형구 내 희귀식물(Korea National Arboretum, 2008)은 CR (멸종위기종: Critically Endangered)의 눈잣나무(Pinus pumila (Pall.) Regel), 이삭단엽란(Microstylis monophyllos (L.) Lindl.) 2분류군, EN (위기종: Endangered Species)의 땃두릅나무(Oplopanax elatus (Nakai) Nakai), 만주송이풀(Pedicularis mandshurica Maxim.) 2분류군, VU(취약종: Vulnerble)의 가문비나무(Picea jezoensis (Siebold & Zucc.) Carrière), 주목(Taxus cuspidata Siebold & Zucc.), 댕댕이나무(Lonicera caerulea var. edulis Turcz. ex Herder), 금강초롱꽃(Hanabusaya asiatica (Nakai) Nakai), 나도옥잠화(Clintonia udensis Trautv. & C.A.Mey.), 자주솜대(Smilacina bicolor Nakai) 6분류군, LC (약관심종: Least Concern)의 개석송(Lycopodium annotinum L.), 세잎종덩굴(Clematis koreana Kom.), 도깨비부채(Rodgersia podophylla A.Gray), 태백제비꽃(Viola albida Palib. ), 만병초(Rhododendron brachycarpum D.Don ex G.Don), 정향나무(Syringa patula var. kamibayshii (Nakai) K.Kim), 꽃개회나무(Syringa wolfii C.K.Schneid.), 과남풀(Gentiana triflora var. japonica (Kusn.) H. Hara), 덩굴꽃마리(Trigonotis icumae (Maxim.) Makino), 금마타리(Patrinia saniculaefolia Hemsl.), 게박쥐나물(Parasenecio adenostyloides (Franch. & Sav. ex Maxim.) H.Koyama), 귀박쥐나물(Parasenecio auriculata (DC.) H.Koyama), 금강애기나리(Streptopus ovalis (Ohwi) F.T.Wang & Y.C.Tang) 13분류군으로 총 24분류군이 확인되었으며, 희귀식물 571분류군의 4.1%에 해당된다.
방형구 내 특산식물(Chung et al., 2017)은 금강초롱꽃, 자주솜대, 진범(Aconitum pseudolaeve Nakai), 청괴불나무(Lonicera subsessilis Rehder), 병꽃나무(Weigela subsessilis (Nakai) L.H.Bailey), 고려엉겅퀴(Cirsium setidens (Dunn) Nakai), 한라사초(Carex erythrobasis H.Lev. & Vaniot) 7분류군으로 확인되었다.
눈측백의 수직적 분포범위는 1,000∼1,600 m로 조사되었으며, 높은 고도에 분포하는 지역은 설악산이며, 가리왕산과 태백산이 가장 낮은 고도인 1,000 m 에서 자생하는 것으로 조사되었다.
설악산의 눈측백은 주로 1,000 m 이상에서 경사가 급하거나 북사면에 발달한 암괴원에 군락을 형성하고 있으며, 1,645∼1,650 m의 북사면에 주로 눈측백이 군락을 이루고 분포한다고 보고하였다(Kwon et al., 2010). 본 조사에서는 기존 보고된 눈측백의 수직적 분포 범위보다 광범위한 수직적 분포로 나타났다. 지형적 특성은 대부분 암석지 또는 전석지이며 낙엽층과 부엽층의 발달이 저조하고 사면경사는 10∼30°로 사면 상부와 능선부에 대부분 자생하는 것으로 조사되었다(Table 1).
Table 1. Environmental factors in study area
군집 분류
흉고직경 1 ㎝ 이상에 대해 매목조사를 실시하여 흉고단면적 분석 결과, 가리왕산이 49.98 ㎡/㏊로 가장 높게 나타났으며, 구성하는 주요 종은 분비나무(Abies nephrolepis (Trautv.) Maxim.) 14.42 ㎡/㏊, 주목 16.03 ㎡/㏊, 사스래나무(Betula ermanii Cham.) 10.14 ㎡/㏊ 등으로 나타났다. 또한 가리왕산에서 눈측백의 흉고단면적이 2.38 ㎡/㏊로 조사지역 중 가장 높게 나타났다.
계방산의 흉고단면적은 20.96 ㎡/㏊으로 나타났으며, 가문비나무 4.73 ㎡/㏊, 사스래나무 10.14 ㎡/㏊, 복장나무(Acer mandshuricum Maxim.) 2.68 ㎡/㏊ 등이 우점하고 있었다.
설악산의 흉고단면적은 11.20 ㎡/㏊로 우점하는 수종은 사스래나무 2.50 ㎡/㏊, 신갈나무(Quercus mongolica Fisch. ex Ledeb.) 1.79 ㎡/㏊, 주목 1.42 ㎡/㏊ 등이 우점하였다.
장산의 흉고단면적은 18.64 ㎡/㏊로 주요 우점하는 수종은 잣나무(Pinus koraiensis Siebold & Zucc.) 4.73 ㎡/㏊, 신갈나무 3.30 ㎡/㏊, 주목 1.39 ㎡/㏊ 등으로 조사되었다.
태백산의 흉고단면적은 33.24 ㎡/㏊로 주요 우점하는 수종은 귀룽나무(Prunus padus L.) 17.07 ㎡/㏊, 분비나무 7.27 ㎡/㏊, 마가목(Sorbus commixta Hedl.) 4.56 ㎡/㏊ 등이 출현하였다.
화악산은 흉고단면적이 3.46 ㎡/㏊으로 조사지역 중 가장 낮게 나타났으며, 주요 수종은 사스래나무 1.37 ㎡/㏊, 분비나무 0.67 ㎡/㏊ 등으로 분석되었다(Table 2).
Table 2. Means of BHA (㎡/㏊) in study area (±SE)
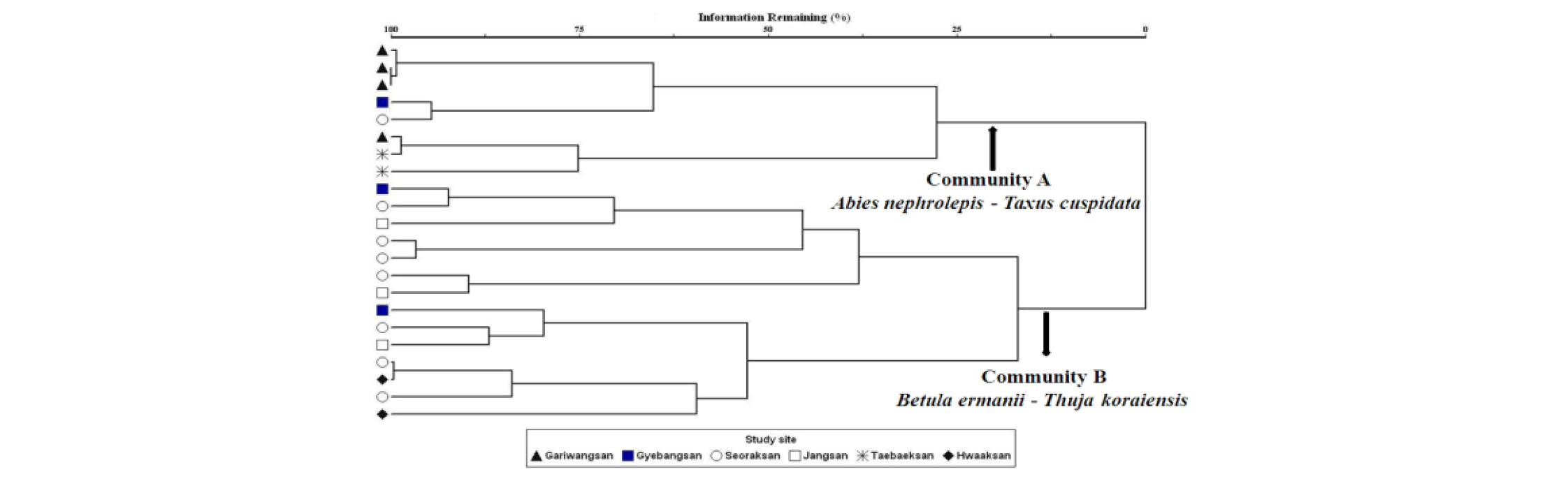
본 조사지역을 바탕으로 군집분류를 실시한 결과 2개 군집으로 분류되었다(Fig. 2). 군집 A (분비나무-주목군집)는 가리왕산(4개), 태백산(2개) 계방산(1개), 설악산(1개)으로 총 8개 방형구가 포함하고 있으며, 분비나무와 주목 등 침엽수종이 우점하고 있는 분비나무-주목군집이다. 또한, 군집 B (사스래나무-눈측백군집)는 계방산(2개소), 설악산(7개소), 장산(3개소), 화악산(2개소)지역으로 총 14개의 방형구를 포함하고 있으며, 활엽수종이 주로 우점하는 것으로 분석되었다. 군집분류 된 두 집단의 목본식생 종조성 차이를 확인하기 위하여 MRPP검정을 실시하였다. 분석결과 두 군집의 종조성의 차이에 대한 통계값이 유의성 있게 나타나 (p < 0.0001, Chance-corrected within- group agreement, A=0.192) 각 군집 종조성의 차이로 인한 적절한 분류로 판단하였다. 일반적으로 군집생태에서 A값은 1에 가까울수록 같은 군집 내 조사구에서 종조성은 동일하다고 할 수 있으며(Mielke and Berry, 1982), A값이 0.192로 분석되어 두 군집이 이질적인 종조성이라고 판단된다.
중요치 분석
상층 식생의 중요치 분석 결과 군집 A (분비나무-주목군집)는 분비나무 17.22%, 주목 16.00%이 우점하는 군집으로 가리왕산, 태백산의 방형구가 포함되어 있다. 군집 B(사스래나무-눈측백군집)는 계방산, 장산, 화악산, 설악산의 방형구를 포함한 사스래나무 14.73%가 우점하는 군집으로 눈측백 9.61%, 잣나무 8.62%, 신갈나무 8.56%가 우점하며, 군집 A에 출현하지 않는 신갈나무, 복장나무, 당단풍나무(Acer pseudosieboldianum (Pax) Kom.), 철쭉(Rhododendron schlippenbachii Maxim.) 등으로 분석되었다(Table 3).
Table 3. Upper layer species importance percentage each community
군집 A에서 생육하고 있는 눈측백의 경우 직립하여 자라는 경향으로 조사되었다. 이러한 생육경향의 원인은 분비나무 등이 우점하는 상층 식생의 피도가 높아 바람의 영향이 적고 임내광량이 충분하지 않아 광 경쟁에 의한 것으로 판단된다. 그러나 군집 B와 같이 관목층의 발달이 높은 지역에서는 눈측백이 포복성으로 자라는 경향으로 나타났다. 이는 군집 B가 군집 A보다 상층 식생의 피도가 상대적으로 낮아 임내 광환경이 양호하여 눈측백을 포함한관목층 내 자원경쟁에 있어 풍부한 광량보다는 토양 내 양분가용을 위한 생육수단에 의하여 포복성의 개체가 많이 나타난 것으로 판단하였다. 또한 군집 B는 낮은 상층 식생의 피도로 바람의 영향에 의하여 눈측백의 수고성장에 방해를 준 것으로 판단하였다.
하층 식생의 중요치 분석결과, 군집 A는 눈측백 11.92%, 분비나무 9.14%의 치수발생이 높았으며, 퍼진고사리(Dryopteris expansa (C.Presl) Fraser-Jenk. & A.C. Jermy) 7.43%, 마가목 4.42%, 주목 3.36%, 관중(Dryopteris crassirhizoma Nakai) 2.76%, 뱀톱(Lycopodium serratum Thunb.) 2.52%, 좀나도히초미(Polystichum braunii (Spenn.) Fée) 2.46%, 애기괭이밥(Oxalis acetosella L.) 2.28%, 만년석송(Lycopodium obscurum L.) 2.13% 등이 우점하고 있으며, 군집 B에 출현하지 않는 찰피나무(Tilia mandshurica Rupr. & Maxim.) 1.34%, 산겨릅나무(Acer tegmentosum Maxim.) 1.18%, 박달나무(Betula schmidtii Regel) 1.18%, 지렁쿠나무(Sambucus sieboldiana var. miquelii (Nakai) Hara) 0.94% 등을 포함하는 군집으로 분석되었다.
군집 B는 눈측백 9.63%, 사스래나무 4.07% 등 치수발생이 높은 것으로 조사되었으며, 퍼진고사리 4.86%, 미역줄나무(Tripterygium regelii Sprague & Takeda) 4.06%, 잣나무 2.88%, 송이풀(Pedicularis resupinata L.) 2.05% 등이 우점하는 것으로 분석되었으며, 군집 A에서는 출현하지 않았던 두루미꽃(Maianthemum bifolium (L.) F.W.Schmidt) 2.49%, 꽃개회나무(Syringa wolfii C.K.Schneid.) 1.70%, 신갈나무 1.61%, 당단풍나무 1.53% 등을 포함하는 군집이다(Table 4).
Table 4. Low layer species importance percentage each community
하층 식생의 중요치를 살펴보면 군집 A는 군집 B에 비해 양치식물류가 높게 분석되었다. 양치식물류는 습윤 한랭한 지역에 주로 생육하는 것으로 보아 군집 A는 군집 B와 비교하여 임분 내 식물종들의 수분스트레스가 적을 것으로 판단된다. 또한, 분비나무, 눈측백 등 상층 식생의 고사로 인하여 군집 A에서 천이 및 임분동태가 유동적인 경향을 보일 것으로 판단되며, 눈측백의 천이과정을 파악하기 위하여 하층 식생의 장기모니터링이 요구된다(Park et al., 2019).
종간 상관관계
식물 군집 내에서 종간 상관관계는 각 식물이 생육하는 환경요인의 이질성을 설명할 수 있는 자료가 된다(Ludwig and Reynolds, 1988). Table 5는 우점하는 상층 식생 15종에 대한 수종간의 상관관계를 분석하였다. 분석결과, 눈측백과 상관관계의 유의성이 인정되는 수종은 분비나무, 주목, 청시닥나무(Acer barbinerve Maxim.), 사스래나무로 나타났다. 그 밖에 신갈나무와 잣나무, 개벚지나무(Prunus maackii Rupr.)와 신갈나무, 귀룽나무와 함박꽃나무(Magnolia sieboldii K.Koch), 당단풍나무와 잣나무, 당단풍나무와 신갈나무, 부게꽃나무(Acer ukurunduense Trautv. & C.A.Mey.)와 마가목(Sorbus commixta Hedlund)이 높은 정의 상관관계를 보였다(p < 0.05).
또한 사스래나무와 주목, 개벚지나무와 잣나무, 마가목과 함박꽃나무, 마가목과 귀룽나무, 청시닥나무와 주목, 청시닥나무와 사스래나무, 복장나무와 가문비나무, 당단풍나무와 개벚지나무, 당단풍나무와 시닥나무(Acer komarovii Pojark.)가 높은 정의 상관관계를 보였다(p < 0.01). Acer 속에 속하는 당단풍나무, 부게꽃나무, 시닥나무는 높은 내한성 수종으로(Cho, 1990) 국내 아고산·고산 생태계에서 주로 우점하고 있는 대표적인 활엽수종으로 본 연구분석에도 Acer속 간 생육환경의 유사한 정도를 확인할 수 있었다.
Table 5. Correlations between all pair-wise combination of major Upper layer species
z(sp.1-Abies nephrolepis (Trautv.) Maxim.; sp.2-Picea jezoensis (Siebold & Zucc.) Carriere; sp.3-Pinus koraiensis Siebold & Zucc.; sp.4-Pinus pumila (Pall.) Regel; sp.5-Thuja koraiensis Nakai; sp.5-Taxus cuspidata Siebold & Zucc.; sp.7-Betula ermanii Cham.; sp.8-Quercus mongolica Fisch. ex Ledeb.; sp.9-Prunus padus L.; sp.10-Sorbus commixta Hedl.; sp.11-Acer barbinerve Maxim.; sp.12-Acer komarovii Pojark.; sp.13-Acer mandshuricum Maxim.; sp.14-Acer pseudosieboldianum (Pax) Kom.; sp.15-Acer ukurunduense Trautv. & C.A.Mey.).
NMS 분석
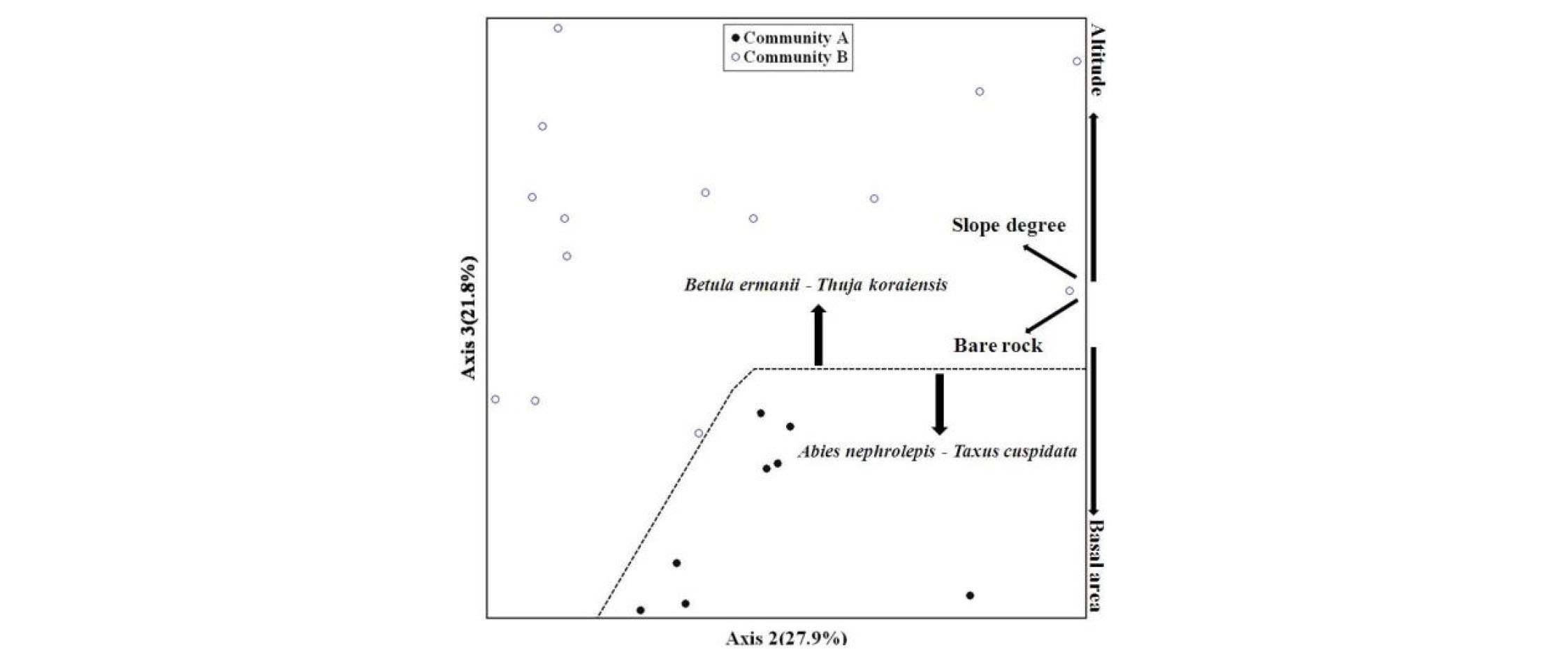
상층 식생의 NMS 분석 결과 높은 설명력을 갖는 2축과 3축을 사용하였으며, 2축의 설명력은 27.9%, 3축의 설명력은 21.8%로 전체 설명력은 49.7%로 분석되었다(Fig. 3). 환경요인은 흉고단면적, 고도, 노암율, 경사가 다른 환경요인에 비해 높은 설명력을 나타났다(cut off R2=0.3). 흉고단면적과 고도는 상반되는 환경요인으로 작용하였다. 군집 A는 상대적으로 흉고단면적이 높으며, 군집 B는 상대적으로 고도와 경사가 높은 수치로 분석되었다. 군집 A의 경우 대경목 단목형태의 침엽수종인 분비나무 및 주목이 다수 생육하기 때문에 나타난 결과로 판단된다. 상층 식생에서 군집 B는 군집 A에 비해 흉고단면적이 낮으며, 참회나무, 두릅나무, 털진달래, 철쭉, 청괴불나무 등 관목성 수종의 비율이 높은 것이 특징이다. 이는 눈측백과 비슷한 층위에 경쟁수종이 많다는 것을 의미한다. NMS를 통한 종조성의 분포역을 살펴보면 두 군집간의 종조성의 분포역이 매우 이질적인 것으로 분석되었다. 상층 식생의 경우 군집 A의 경우 군집 B에 비하여 좁은 종조성의 분포를 나타내고 있어 군집 B의 종조성이 군집 A에 비하여 광범위하고 복잡한 종조성으로 나타난 것으로 판단하였다.
눈측백 자생지 내 하층 식생의 NMS 분석결과 Fig. 4와 같다.좌측은 하층 식생 조사지의 종조성 분포이며, 우측은 하층 식생에 나타난 식물종의 분포를 나타낸 결과이다. 1축의 설명력은 27.0%, 2축의 설명력은 25.4%로 전체 설명력은 42.4%로 나타났으며, 종조성 및 식물종의 분포에 영향을 미치는 주요 환경요인은 상층 및 하층 식생의 종풍부도와 낙엽층 깊이가 영향을 미치는 것으로 분석되었다(cut off R2=0.25). 눈측백 개체군 내 종조성의 분포는 상층 식생에 비하여 군집 A와 B에서는 뚜렷한 차이가 나타나지는 않았다. 군집 B가 군집 A보다 종조성의 다양성 및 분포영역이 광범위하여 상층 식생의 경향과 유사한 결과로 나타났다. 하층 식생에 자생하는 주요 식물종의 분포를 살펴보면 는쟁이냉이, 가문비나무, 도깨비부채, 산겨릅나무, 진달래, 개다래, 털개회나무, 병꽃나무 등은 타 식물종과 상대적으로 상층 및 하층 식생의 종풍부도와 낙엽층 깊이가 낮은 곳에 생육하는 종으로 분석되었다. 상층과 하층 식생의 종풍부도가 높고 낙엽층 깊이가 깊은 곳에 생육하는 종은 좀나도히초미, 바늘사초, 두루미꽃, 철쭉으로 분석되었다. 단풍취, 청괴불나무, 큰꼭두서니, 선포아풀, 생열귀나무, 산구절초, 산새풀, 송이풀은 상층과 하층 식생의 종풍부도는 높으나 낙엽층깊이가 낮은 곳에 주로 생육하는 종으로 분석되었다.
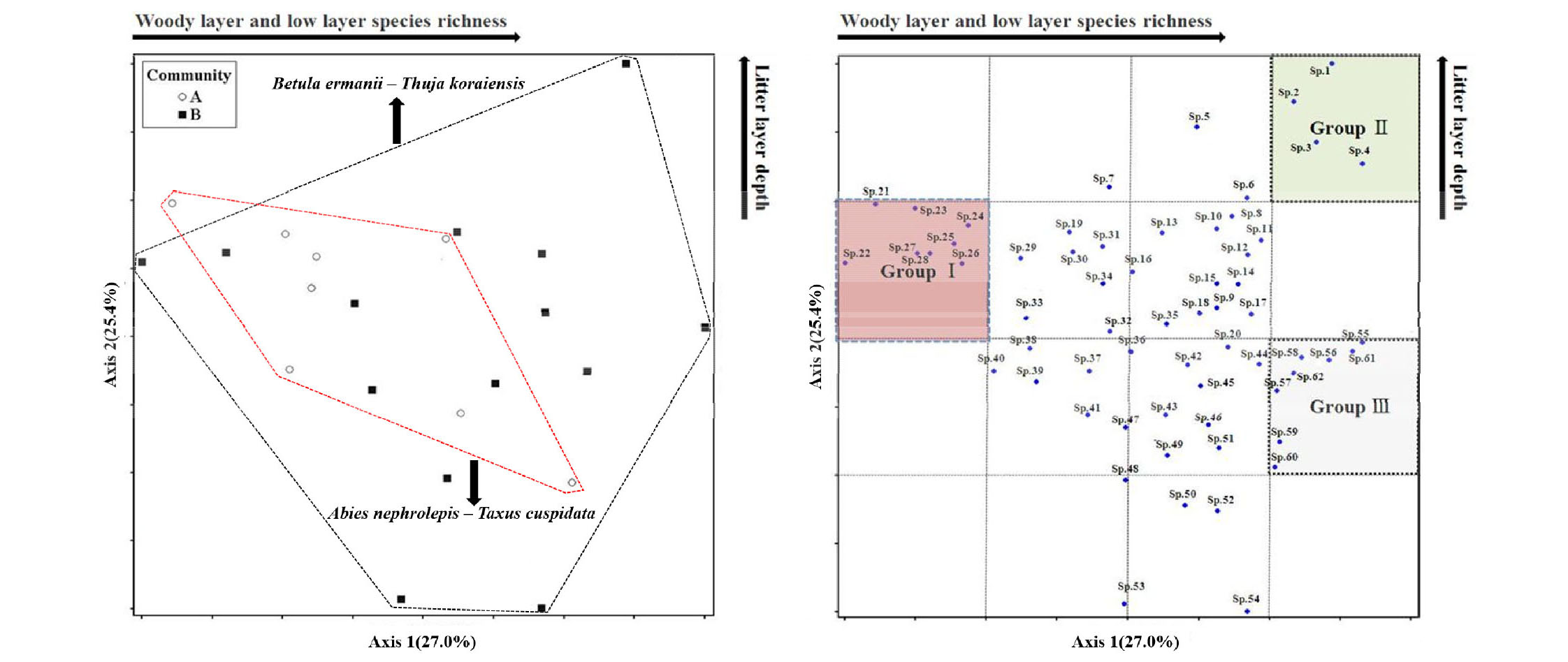
Fig. 4.
NMS distribution of study area(Low layer species, cut off R2=0.25). *minor species was omitted : sp.1-Polystichum braunii; sp.2-Dryopteris expansa; sp.3-Maianthemum bifolium; sp.4-Oxalis acetosella; sp.5-Deparia pycnosora; sp.6-Parasenecio adenostyloidesa; sp.7-Athyrium brevifrons; sp.8-Sasa borealis; sp.9-Magnolia sieboldii; sp.10-Pinus koraiensis; sp.11-Pseudostellaria heterophylla; sp.12-Rhododendron mucronulatum; sp.13-Cirsium setidens; sp.14-Clintonia udensis; sp.15-Acer pseudosieboldianum; sp.16-Acer komarovii; sp.17-Actinidia polygama; sp.18-Taxus cuspidata; sp.19-Tilia mandshurica; sp.20-Sedum polytrichoides; sp.21-Cardamine komarovii; sp.22-Picea jezoensis; sp.23-Rodgersia podophylla; sp.24-Acer tegmentosum; sp.25-Actaea asiatica; sp.26-Polystichum tripteron; sp.27-Trigonotis icumae; sp.28-Hydrangea serrata f. acuminata; sp.29-Syringa reticulata var. mandshurica; sp.30-Paris verticillata; sp.31-Thuja koraiensis; sp.32-Acer barbinerve; sp.33-Sorbaria sorbifolia var. stellipila; sp.34-Rhododendron schlippenbachii; sp.35-Rhododendron brachycarpum; sp.36-Carex onoei; sp.37-Abies nephrolepis; sp.38-Betula ermanii; sp.39-Primula jesoana; sp.40-Lycopodium annotinum; sp.41-Crepidiastrum chelidoniifolium; sp.42-Patrinia saniculaefolia; sp.43-Parasenecio auriculata; sp.44-Prunus maackii; sp.45-Isodon inflexus; sp.46-Prunus padus; sp.47-Euonymus macropterus; sp.48-Astilbe rubra; sp.49-Trigonotis icumae; sp.50-Festuca ovina; sp.51-Asplenium incisum; sp.52-Serratula coronata var. insularis; sp.53-Pinus pumila; sp.54-Syringa wolfii; sp.55-Ainsliaea acerifolia; sp.56-Lonicera subsessilis; sp.57-Rubia chinensis; sp.58-Tripterygium regelii; sp.59-Rosa davurica; sp.60-Dendranthema zawadskii; sp.61-Calamagrostis langsdorfii; sp.62-Pedicularis resupinata.
지표종 분석
지표종 분석은 확률화 검정(randomization test)을 통한 각 군집 내 종들에 대한 지표값을 나타내며, 높은 값의 지표종은 군집간의 오차가 거의 없는 현재 지역에서 현존하는 종(Dufrene and Legendre, 1997)이다.
지표종 분석결과, 유의차(p < 0.05) 있는 지표종 분석은 상층 식생에서 3분류군, 하층 식생에서 6분류군이 분석되었다(Table 6). 상층 식생에서 분비나무, 주목, 귀룽나무가 지표종으로 분석되었으며, 하층 식생에서 좀나도히초미, 만년석송, 애기괭이밥, 찰피나무, 산겨릅나무, 주목 등 6분류군으로 총 8분류군이 지표종으로 분석되었다. 분석된 8분류군의 지표종은 눈측백 자생지 보전과 복원에서 우선적으로 고려되어야 할 것으로 판단된다.
Table 6. Indicator species in study area
고찰
본 연구는 눈측백 자생지의 식생과 환경요인을 분석하여 향후 보전과 관리를 위한 기초자료를 제시하고자 수행되었다. 눈측백은 해발고도 1,000∼1,588 m에 자생하고 있으며, 경사는 10∼30°로 대부분 북사면에 군락을 이루고 있었다. 6개 지역 내 22개 방형구에서 150분류군의 관속식물이 확인되었다. 눈측백 자생지의 흉고단면적은 가리왕산지역이 49.98 ㎡/㏊로 가장 높게 나타났으며, 화악산지역이 3.46 ㎡/㏊으로 가장 낮게 나타났다. 군집분석에 의해 2개의 군집(분비나무-주목군집, 사스래나무-눈측백 군집)으로 구분되었으며, MRPP 검정 결과 군집은 유의성 있게 구별되는 것으로 나타났다. 지표종 분석은 분비나무-주목군집에서만 나타났으며, 유의한 종으로는 분비나무, 주목, 귀룽나무, 좀나도히초미, 만년석송, 애기괭이밥 등 총 8분류군으로 분석되었다.
최근 국내에서 지구환경변화로 인한 눈측백의 분포역 변화를 예상하는 선행연구를 바탕으로(Kong, 2005), 본 연구를 통하여 눈측백 개체군 고사가 다수 확인되어 눈측백의 현지내·외 보전방안이 요구된다. 현지내 보전(in situ)을 위한 활동은 보호구역을 설정하여 보호 및 보전대책을 강구하는 방법이 있다. 멸종위기에 처한 식물에 대한 보전대책으로 IUCN은 생물다양성 보전을 위한 현지내 보전전략으로 보호구역을 설정을 제안하고 있으며(IUCN, 2020), 국제보전협회(CI: Conservation International)에서는 생물다양성 핵심지역(Biodiversity Hotspot)으로 지정 및 보전되고 있다.
대부분의 눈측백 자생지는 국립공원, 군사보호지역, 도립공원, 산림유전자원보호구역 등 보호지역으로 지정되어 관리되고 있으나 장산은 산 정상부만 보호구역으로 국한되어 눈측백을 포함한 희귀식물 보전을 위해서는 보호구역 지정면적을 확대할 필요가 있다고 판단된다.
또한, 눈측백 개체군이 자생한 분비나무-주목이 우점하였던 군집 A의 식생유형은 남한 내 유적군락(remnant vegetation)으로 식생학적 가치가 높으며, 습윤 침엽수림 지역에서서 제한적으로 존재하는 남한 내 희귀식생 유형으로 판단된다. 또한, 눈측백이 수고 10 m 이상의 개체를 다수로 확인하였다. 하지만 직립형 개체목의 고사현상이 두드러져 눈측백의 개체군 동태파악을 위한 지속적인 모니터링이 필요할 것으로 판단되었다.
군집 B에서는 직립형 눈측백이 1 m 이상으로 생육하고 있었으나, 대부분 고사하였으나 주변으로 치수가 발생하여 포복성으로 자생하고 있었다. 눈측백 치수는 수관열림 등에 따른 광환경, 수분스트레스, 타 관목류(참회나무, 두릅나무, 털진달래, 철쭉)간 종간 경쟁에 약한 것으로 판단되었다. 따라서 눈측백 집단 내 쇠퇴 원인과 우선적 보존지역을 설정하기 위하여 눈측백 개체군의 유전자 분석이 필요할 것으로 판단된다(Milligan et al., 1994).
눈측백 자생지를 대상으로 현지내 보전(in situ)만으로는 한계가 있으며, 현지외 보전(ex situ)도 동시에 추진되어 할 것으로 판단된다. 현지외 보전원은 생체 수집으로 보존원 조성, 종자은행(seed bank) 및 채종원 구축을 통하여 눈측백 개체군의 보전 프로세스를 구체적으로 설정한 보전전략이 필요하다. 향후 눈측백 등 개체군에 대한 보전방안이 안정적으로 유지된 후, 눈측백 자생지의 산림 생태계 전반에 대한 보전대책이 수립되어야 할 것으로 판단된다.
적요
본 연구는 눈측백 자생지의 식생과 환경요인을 분석하여 향후 보전과 관리를 위한 기초자료를 제시하고자 수행되었다. 눈측백은 해발고도 1,000∼1,588 m에 자생하고 있으며, 경사는 10∼30°로 대부분 북사면에 군락을 이루고 있었다. 6개 지역 내 22개 방형구에서 150분류군의 관속식물이 확인되었다. 눈측백자생지의 흉고단면적은 가리왕산지역이 49.98 ㎡/㏊로 가장 높게 나타났으며, 화악산지역이 3.46 ㎡/㏊으로 가장 낮게 나타났다. 군집분석에 의해 2개의 군집(분비나무-주목군집, 사스래나무-눈측백 군집)으로 구분되었으며, MRPP 검정 결과 군집은 유의성 있게 구별되는 것으로 나타났다. 지표종 분석은 분비나무-주목군집에서만 나타났으며, 유의한 종으로는 분비나무, 주목, 귀룽나무, 좀나도히초미, 만년석송, 애기괭이밥 등 총 8 분류군으로 분석되었다.
