

서 언
재료 및 방법
샘플 수집
Genomic DNA 추출
엽록체 유전체 조립 및 계통수 분석
변이지역 분석
결 과
엽록체 유전체 조립
엽록체 유전체 구조 분석
유전자 주석 결과
계통수 분석 결과
변이지역 분석 결과
고 찰
적 요
서 언
돌나물과(Crassulaceae)의 기린초속(Phedimus)은 약 20여 종의 다육식물로 구성되며, 주로 유럽과 아시아 전역, 특히 한국, 중국, 일본 등지에 분포하고 있다(Lee, 2023; Thiede and Eggli, 2007). 분류학적으로 기린초속은 과거 다계통군(polyphyletic group)으로 밝혀진 돌나물속(Sedum)의 일부분으로 분류되었으나, 이후 별도의 속으로 독립되었다(Moon and Jang, 2020; Ohba et al., 2000). 이 속에 속하는 식물은 주로 다년생 초본이지만, 일부는 드물게 일년생으로도 나타나며, 다육질의 줄기와 협소한 기부를 가지는 잎이 특징적이다. 또한, 대부분의 종은 잎이 어긋나게 배열되고, 잎 가장자리에 작은 포엽이 형성되는 경우가 많으며, 줄기는 때때로 목질화되기도 한다(Moon and Jang, 2020). 꽃차례는 주로 정생 총상(terminal raceme) 구조를 가지며, 일반적으로 5개의 꽃덮이(tepals), 5개의 심피(carpels), 그리고 심피에 부착된 두 배수의 수술(stamens)로 구성된 노란색 꽃을 갖는다(Ohba et al., 2000; Rafinesque, 1817; 't Hart and Bleij, 2003). 이들은 주로 암석지대에 서식하며, 높은 내건성과 낮은 수분 요구량을 특징으로 한다(Lee et al., 2021).
속리기린초(P. zokuriensis)와 태백기린초(P. latiovalifolium)는 한국 자생종으로 속리산, 함백산과 같은 일부 한정된 지역에서 분포하며(Park et al., 2020; Yoo and Park, 2016), 속리기린초(P. zokuriensis)의 경우 포복형이 존재하고 잎이 난형 또는 타원형으로 존재하고, 잎 가장자리가 매끈하고 톱니가 약하게 자라며 마주나거나 어긋나기로 배열된다. 꽃은 노란색 별모양의 작은 꽃을 피우며, 산방상 또는 취산상으로 피며, 꽃대 없이 바로 달린다(Moon and Jang, 2020). 태백기린초(P. latiovalifolium)의 경우 포복형이 없으며, 단간성 직립 줄기로 줄기 끝에 로제트(rosette)를 형성한다. 잎은 넓은 난형이고 두꺼운 다육질로 잎 가장자리에 잔톱니가 특징적이다. 꽃은 노란색 별모양으로 복산형 구조를 형성한다(Yoo and Park, 2016). 한국 전통의학에서 기린초속 종들은 지혈 및 해독 작용으로 널리 사용되어 왔다(Park et al., 2008). 이들은 오랜 기간 혈액 순환 촉진, 지혈, 항염 작용 등의 다양한 약리적 효과로 전통적으로 활용되어 왔다(Park et al., 2008; Yoon et al., 2020). 최근 연구에서는 이들 식물의 다양한 부위에서 강력한 항산화 활성이 확인되어 피부 보호, 미백 효과 등 기능성 화장품 원료로서의 잠재력이 주목받고 있다(Kim et al., 2023; Yoon et al., 2020). 또한, 기린초속 추출물은 항균 및 항진균에도 효과를 나타내는 것으로 보고되었다(Lee et al., 2024a; Wang et al., 2024a).
아울러, 최근 국내에서 보고된 식물상 조사(Kwon, 2025), 산림생태복원 모니터링(Lee et al., 2024b), 및 관련 연구들(Hwang et al., 2023; Yi et al., 2023)은 자생 식물의 분포와 다양성 연구의 지속성을 보여주며, 본 연구와 같이 특정 속에 대한 분자계통학적 접근과 상호 보완될 수 있다. 또한 엽록체 유전체 기반의 분자 마커 개발(Lee et al., 2023)과 형태-분자 통합 계통학적 연구(Yu et al., 2023) 역시 본 연구의 맥락을 뒷받침한다. 한편, 형태적 특징이 유사한 종들 사이에서는 전통적인 분류학적 접근만으로는 정확한 동정이 어려운 경우가 많다. 특히 한국 자생 기린초속은 고산지대 등 제한된 서식지에 분포하나 형태적 차이는 뚜렷하지 않은 편이기 때문에 분자계통학적 접근을 통한 재검토가 요구된다(Moon and Jang, 2020). 이러한 관점에서 엽록체 유전체(chloroplast genome) 서열은 고유한 유전자 서열을 비교함으로써 종의 신속하고 정확한 동정을 가능하게 하는 Insertion/Deletion (InDel), Single Nucleotide Polymorphism (SNP), Simple Sequence Repeat (SSR)와 같은 DNA 마커(marker)의 제공 source로서 점차 중요한 역할을 하고 있다(Li et al., 2020; Song et al., 2022; Wang et al., 2024b; Zhu et al., 2023).
본 연구에서는 기린초속(Phedimus) 두 종의 완전한 엽록체 유전체를 시퀀싱하여 조립 및 유전자 지역을 주석화하였으며, 이들 사이의 계통발생적 관계를 명확히 하고자 계통유전학적 분석을 수행하였다. 아울러 엽록체 유전체 염기서열이 향후 마커개발을 위한 source로 사용가능한지 여부를 보기 위해 두 종간의 후보 변이지역을 탐색하였다. 본 연구의 결과는 향후 기린초속(Phedimus) 식물의 유전적 다양성 및 보전 연구에 중요한 기초 자료로 활용될 수 있을 것이다.
재료 및 방법
샘플 수집
속리기린초(P. zokuriensis, 유전자원번호: IT317349) 식물체(Fig. 1A)는 속리산(36°33'30" N, 127°51'20" E)에서, 태백기린초(P. latiovalifolium, 유전자원번호: IT317342) 식물체(Fig. 1B)는 함백산(37°08'50" N, 128°54'10" E)에서 수집되었다. 수집된 표본은 각각 고유 수집번호 SYCS102 (P. zokuriensis) 및 SYCS103 (P. latiovalifolium)로 기재되어 삼육대학교 자연과학연구소(담당자: 남상용 교수, namsy@syu.ac.kr)에 영구적으로 보존되었다.

Genomic DNA 추출
두 종의 잎 조직으로부터 유전체 DNA는 수정된 CTAB (cetrimonium bromide) 기반의 방법을 이용하여 추출하였으며(Kuo et al., 2022), 추출된 DNA는 Nanodrop (Thermo Scientific Inc., Waltham, MA, USA)과 아가로스 겔 전기영동을 통해 농도 및 순도(무결성)를 검증하였다. Illumina paired-end 유전체 라이브러리는 NEXTflex® Rapid DNA sequencing kit (Bioo Scientific, Austin, TX, USA)를 사용하여 제조사의 매뉴얼에 따라 제작하였다.
엽록체 유전체 조립 및 계통수 분석
엽록체 유전체는 NovaSeq 6000 (Illumina Inc., San Diego, CA, USA) 플랫폼을 통해 시퀀싱하였으며, 시퀀싱된 원시 데이터는 Trimmomatic (Bolger et al., 2014)을 사용하여 저품질 및 어댑터 서열을 제거하여 고품질의 Paired-end (PE) 리드(read)를 확보하였다. 확보된 고품질 PE 리드는 NOVOplasty ver. 4.3.1 (Dierckxsens et al., 2017)을 사용하여 엽록체 유전체를 조립하였으며, 이후 PE 리드 매핑(Kim et al., 2015)을 통한 수작업 교정을 통해 최종 유전체 서열을 확정하였다. 엽록체 유전체의 주석(annotation)은 GeSeq (Tillich et al., 2017)을 통해 수행되었으며, 필요시 수작업을 통해 추가 교정하였다.
완전한 엽록체 유전체 서열은 각각 P. zokuriensis (GenBank ID: OP452906)와 P. latiovalifolium (GenBank ID: OP452907)으로서 GenBank에 제출되었다. 이들의 BioProject, BioSample, SRA정보는 다음과 같다. P. zokuriensis의 경우 BioProject PRJNA881022, BioSample SAMN30881463, SRA SRR21601458로 등록되어 있고, P. latiovalifolium의 경우 BioProject PRJNA881036, BioSample SAMN30884597, SRA SRR21733744로 등록되어 있다.
속리기린초(P. zokuriensis) 및 태백기린초(P. latiovalifolium)의 계통발생적 위치를 분석하기 위해 기린초속(Phedimus) 및 돌나물속(Sedum)의 20종과 외군(outgroup)으로 선택된 Meterostachys sikokianus의 엽록체 유전체를 MAFFT version 7.526 (Katoh et al., 2002)을 사용하여 정렬하였다. 정렬된 엽록체 유전체 서열을 기반으로 MEGA version 11.0.13 (Tamura et al., 2021)을 사용하여 maximum likelihood (ML)을 통한 계통수(phylogenetic tree)를 bootstrap 1,000회로 구축하였다.
변이지역 분석
엽록체 유전체 간의 변이 분석은 MUMmer version 3 패키지(Kurtz et al., 2004)를 사용하여 수행되었다. 엽록체 유전체 서열은 기본 설정값으로 nucmer를 이용해 정렬되었으며, 생성된 정렬 파일(.delta)은 show-snps명령어에 -ClrT옵션을 적용하여 단일 염기 다형성(SNP) 및 삽입/결실 변이(InDel)를 추출하는 데 사용되었다. 변이들은 기준(reference) 또는 질의(query) 서열 중 어느 한 쪽에 공백(gap)이 존재하는지 여부에 따라 SNP와 indel로 추가 분류되었다. 또한 SSR 분석을 위해 misa.pl (https://github.com/cfljam/SSR_marker_design/ blob/master/misa.pl)이 사용되었으며, mono, di, tri, tetra, penta, hexa-mer 범위 기준으로 탐색하였다.
결 과
엽록체 유전체 조립
속리기린초(P. zokuriensis)의 완전한 엽록체 유전체(OP45 2906)는 총 151,728 bp의 길이를 가지며, G+C 함량은 37.74%로 나타났다. 시퀀싱된 리드는 P. zokuriensis 엽록체 유전체 서열에 매핑된 결과, 평균 1,570배의 높은 커버리지 깊이를 보였다(Fig. 2A). 한편, 태백기린초(P. latiovalifolium)의 완전한 엽록체 유전체(OP452907)는 151,796 bp의 길이와 37.72%의 G+C 함량을 가지며, 매핑된 리드의 평균 커버리지 깊이는 2,481배로 확인되었다(Fig. 2B).
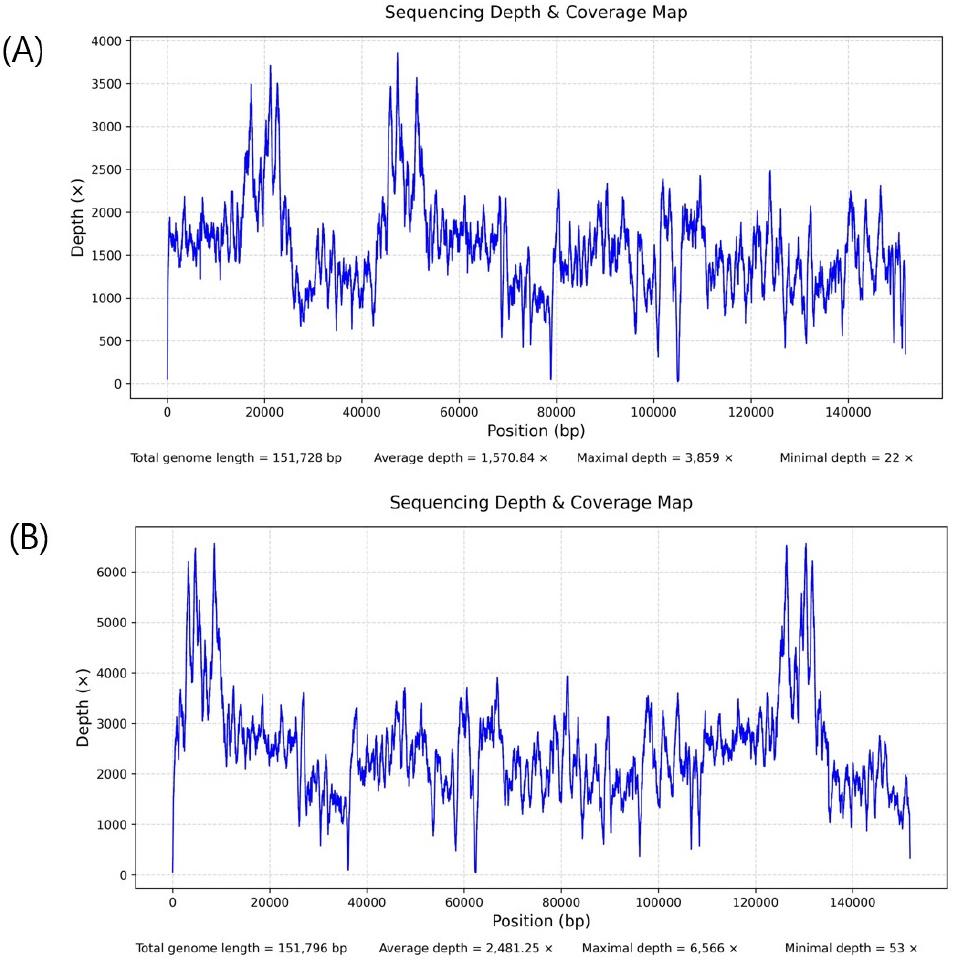
Fig. 2.
Chloroplast genome coverage depth maps of (A) P. zokuriensis and (B) P. latiovalifolium. For P. zokuriensis, the maximum coverage depth is 3,859×, the minimum is 22×, and the average is 1,570×. For P. latiovalifolium, the maximum coverage depth is 6,566×, the minimum is 53×, and the average is 2,481×.
엽록체 유전체 구조 분석
P. zokuriensis의 엽록체 유전체(OP452906)는 83,067 bp의 대형 단일 복제 영역(LSC), 16,697 bp의 소형 단일 복제 영역(SSC), 그리고 25,982 bp의 반복 역위 구간(IRa 및 IRb)으로 구성되어 있다(Fig. 3A). 반면, P. latiovalifolium의 엽록체 유전체(OP452907)는 83,133 bp의 LSC영역, 16,699 bp의 SSC영역, 그리고 각각 25,982 bp의 IRa 및 IRb구간으로 이루어져 있다(Fig. 3B).
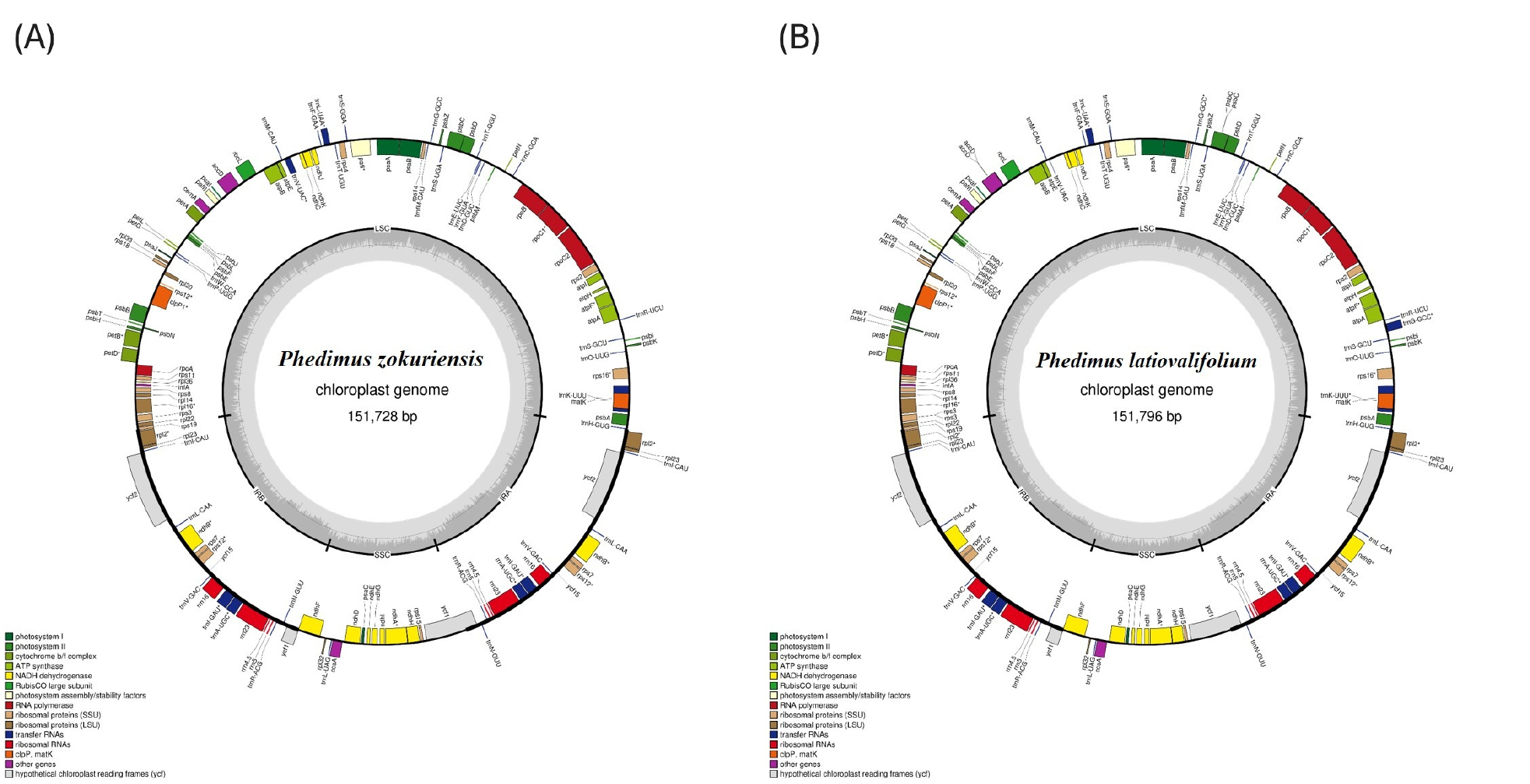
Fig. 3.
Circular maps of the chloroplast genomes of (A) P. zokuriensis and (B) P. latiovalifolium. Genes located inside the circle are transcribed in the counterclockwise direction, while those outside are transcribed in the clockwise direction. Gene colors indicate functional categories. The large single-copy (LSC), small single-copy (SSC), and inverted repeat (IRa and IRb) regions are also indicated.
유전자 주석 결과
속리기린초(P. zokuriensis)의 엽록체 유전체는 총 132개의 유전자를 포함하고 있으며, 이 중 88개는 단백질 코딩 유전자, 35개는 tRNA 유전자, 8개는 rRNA 유전자로 구성되어 있다. 이러한 132개의 예측된 유전자는 19개의 기능 범주로 분류되었으며, 이는 Table 1에 제시되어 있다. 이들 중 gene family를 구성하지 않는 single-copy 형태로 존재하는 유전자는 accD, ccsA, cemA, clpP1, infA, matK, rbcL 7개이며, 나머지 유전자는 모두 multi-copy 형태로 존재한다(Table 1). 반면, 태백기린초(P. latiovalifolium)의 엽록체 유전체는 총 133개의 유전자를 포함하고 있으며, 이 중 88개는 단백질 코딩 유전자, 37개는 tRNA 유전자, 8개는 rRNA 유전자로 구성되어 있다. 이러한 133개의 예측된 유전자는 19개의 기능 범주로 분류되었으며, 이는 Table 2에 제시되어 있다. 이들 중 accD, ccsA, cemA, clpP1, infA, matK, rbcL의 7개 유전자는 single-copy 형태로 존재하는 것으로 확인되었으며, 나머지 유전자는 multi-copy 형태로 존재한다(Table 2).
Table 1.
Gene prediction of the chloroplast genome of P. zokuriensis.
Table 2.
Gene prediction of the chloroplast genome of P. latiovalifolium.
계통수 분석 결과
본 계통수는 Phedimus 및 Sedum 속 총 20개 종의 엽록체 유전체 서열을 기반으로 maximum likelihood (ML) 기반으로 구축하였다. 엽록체 유전체 서열은 GenBank에서 수집되었으며, Meterostachys sikokianus를 외군(outgroup)으로 설정하였다. 계통학적 분석 결과 2개의 단일계통군이 발견되었으며, 1개의 계통군은 Phedimus 속에 속하는 14종(P. takesimensis 및 P. takesimense는 동일한 종으로 간주), 다른 한 개의 계통군은 Sedum 속에 속하는 8종으로, bootstrap 값 1,000회 반복 분석을 통해 두 군 모두 99% 이상의 bootstrap 값을 얻었으며 속리기린초(P. zokuriensis) 및 태백기린초(P. latiovalifolium) 모두 Sedum 속과 확연히 분리되는 Phedimus 속에 포함됨을 보였다(Fig. 4).
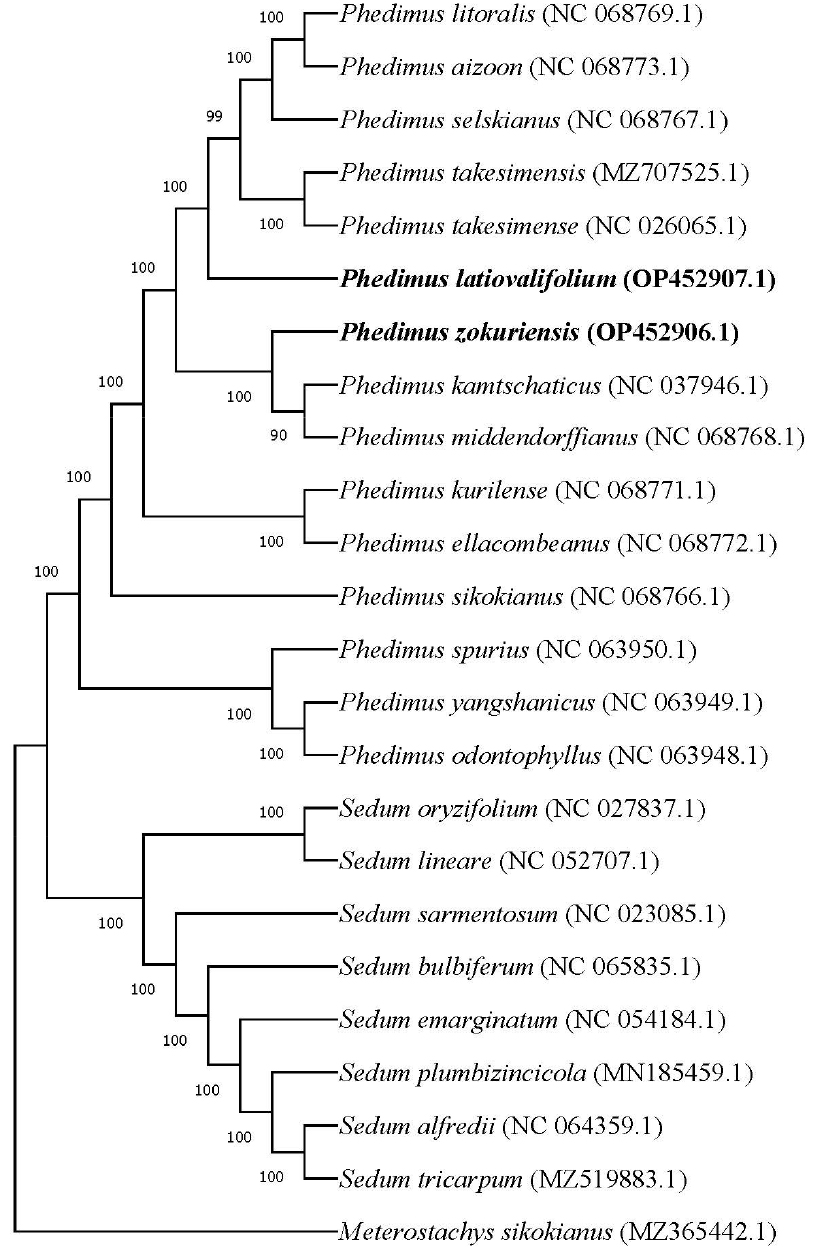
Fig. 4.
Maximum-likelihood phylogenetic tree showing the relationships between P. zokuriensis and P. latiovalifolium (highlighted in bold) and their close relatives. The analysis was based on complete chloroplast genome sequences from P. zokuriensis, P. latiovalifolium, and 20 other species within the Crassulaceae (with P. takesimensis and P. takesimense treated as the same species), using Meterostachys sikokianus as an outgroup. Bootstrap support values based on 1,000 replicates are indicated at each node.
변이지역 분석 결과
속리기린초(P. zokuriensis)와 태백기린초(P. latiovalifolium) 엽록체 염기서열 간 변이지역 탐색을 위해 SSR 분석을 수행한 결과 두 종의 차이를 보이는 SSR 변이지역은 발견되지 않았다. 반면 InDel 및 SNP 변이지역 탐색 결과 총 57개 indel의 후보 변이지역 및 총 168개 SNP의 후보 변이지역을 LSC, SSC, IR 지역에서 각각 확인하였다(Fig. 5). 변이 위치 분석 결과 IR 지역에서는 차이를 보이는 indel 및 SNP 모두 발견되지 않았으나 LSC 지역에서는 53개의 indel, 136개의 SNP 지역이 발견되었고, SSC 지역에서는 4개의 InDel, 32개의 SNP 지역이 발견되었다.
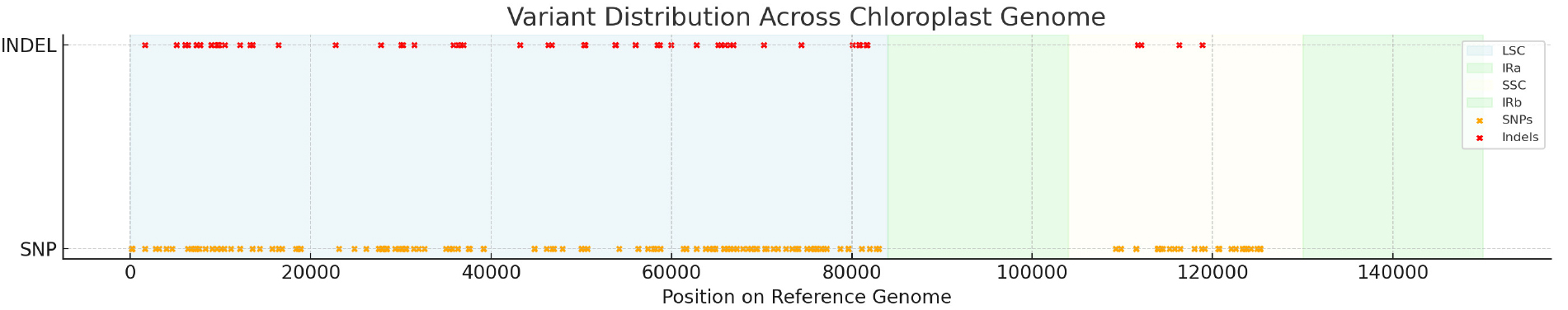
Fig. 5.
Distribution of sequence variants across the chloroplast genome of P. zokuriensis and P. latiovalifolium. SNPs and indels between P. zokuriensis and P. latiovalifolium were identified using the complete chloroplast genome of P. latiovalifolium as the reference. Variants are plotted along the genome coordinate axis, with SNPs shown in orange and indels in red. The structural regions of the chloroplast genome, LSC, SSC, and IR regions, are indicated by semi-transparent background shading.
고 찰
본 연구에서는 대한민국 속리산에서 채집한 속리기린초(P. zokuriensis)와 함백산에서 채집한 태백기린초(P. latiovalifolium)의 엽록체 유전체를 해독 및 비교하여 두 종의 분류학적 위치와 유연관계를 규명하였다. 분석 결과, 두 종의 엽록체 유전체 크기와 구조는 전반적으로 유사했으며, 88개의 단백질 코딩 유전자, 35개의 tRNA 유전자, 8개의 rRNA 유전자가 전반적으로 유사하였으나(P. latiovalifolium에서 tRNA 2개 증가 등) 일부 차이가 확인되었다. 다만, 태백기린초에서 추가로 확인된 2가지 tRNA 유전자의 개수 차이, 즉 trnG-GCC의 복제수 차이(1 copy vs. 2 copies) 및 trnV-UAC의 존재 여부(0 copy vs. 1 copy)는 종 간 차이를 나타내었으며, 이러한 변이 영역은 향후 두 종의 종 경계 규명에 활용 가능한 분자 표지로 기능할 수 있을 것으로 판단된다(Dong et al., 2015; Shaw et al., 2014).
계통학적 분석은 속리기린초(P. zokuriensis)및 태백기린초(P. latiovalifolium)를 포함한 Phedimus 종에 대한 엽록체 유전체 기반의 유연관계를 규명하기 위해 수행되었으며, 이를 통해 엽록체 염기서열이 확보된 두 종의 분류학적 위치를 명확히 하고자 진행되었다. 계통학적 분석 결과, 두 개의 단일계통군이 발견되었으며, 하나는 돌나물속(Sedum)에 속하는 8종, 다른 하나는 기린초속(Phedimus)에 속하는 14종으로, 두 군 모두 99% 이상의 bootstrap 값을 통해 지지받았다(Fig. 4). 본 연구에서 진행된 계통학적 분석에서 속리기린초(P. zokuriensis, OP452906.1)및태백기린초(P. latiovalifolium, OP452907.1) 두 종 모두 Phedimus 속에 위치하며, Sedum 속과는 뚜렷한 구별을 보였다. Phedimus 속은 과거 Sedum 속에 포함되어 있었으나 현재는 분자생물학적 연구 결과에 따라 서로 독립적인 속으로 인식되고 있다(Moon and Jang, 2020). 분석 결과, Phedimus 속은 bootstrap 값 90 이상의 단일 clade로 나타났다. P. latiovalifolium은 P. takesimensis, P. selskianus, P. aizoon, P. litoralis 등과 강하게 지지되는 clade를 형성하며, 이들과 유전적으로 가까운 관계를 가진다. 반면, P. zokuriensis는 P. kamtschaticus 및 P. middendorffianus와 함께 별도의 clade에 속하였다. 이는 두 종이 형태적, 생태적, 지리적 요인에 의해 독립적으로 분화했을 가능성을 시사한다(Rundle and Nosil, 2005; Schluter, 2001). 따라서 본 연구 결과는 속리기린초(P. zokuriensis)와 태백기린초(P. latiovalifolium)가 독립된 종으로 분화되었음을 보여주는 계통학적 근거를 제공한다.
그러나 이러한 유전적 근접성을 생물학적-생태학적 맥락에서 심층적으로 이해하기 위해 다음과 같은 보완적 논의가 필요하다. 첫째, 유전체 수준의 유사성은 확인되었으나 두 종의 형태적 특징과 생태적 적응 차이에 대한 정보는 아직 부족하다. 속리기린초와 태백기린초는 각각 속리산과 함백산이라는 서로 다른 고도와 지형 조건의 서식지에서 자생하므로 생장 습성, 잎과 꽃의 구조, 생육 시기 등에서 차이를 보일 가능성이 크다(Körner, 2007). 추후에 이러한 형태적 형질과 생태적 틈새(niche differentiation)를 체계적으로 비교하면 엽록체 유전체 기반 계통 결과와의 상관성을 더욱 정밀하게 해석할 수 있을 것이다(Dayrat, 2005). 둘째, 두 종의 계통적 유사성이 시사하는 분화의 진화적 배경에 대한 논의가 필요하다. 속리산(중부 내륙)과 함백산(동부 고산)은 지리적으로 떨어져 있으며, 고도 차이에 따른 환경 이질성과 빙하기 이후의 식물 분포 변화가 종 분화를 촉진했을 가능성이 있다(Hewitt, 2000). 특히, 상대적으로 고산 환경에 적응한 P. latiovalifolium에 비해(Park et al., 2020), 비교적 중간지대와 저지대 기후에 특화된 P. zokuriensis와는 다른 진화 경로를 거쳤을 수 있으며, 이러한 지리적-생태적 분리는 두 종 간 유전적 분화를 설명하는 핵심 요인이 될 수 있다(Qiu et al., 2011). 셋째, 본 연구는 엽록체 유전체(모계성 유전) 서열 정보만을 활용하였기에, 부계 또는 양계성 유전 정보를 포함하는 핵유전체 자료는 반영하지 못하였다. 이는 과거 잡종화나 유전자 유입(introgression)의 가능성이 존재하는 분류군에서 계통 해석의 정밀도를 제한할 수 있으므로(Rieseberg and Soltis, 1991), 향후 연구에서는 MIG-seq, RAD-seq, 또는 전장 핵유전체 분석과 같은 다면적 접근법을 적용하여 유전체 전반의 정보를 확보할 필요가 있다(Peterson et al., 2012; Suyama and Matsuki, 2015). 이러한 통합적 분석은 형태적, 생태적 특성과 유전 정보를 결합해 두 종의 진화사 및 종 경계에 대한 보다 정교하고 종합적인 이해를 가능하게 할 것이다(Dayrat, 2005).
적 요
본 연구에서는 한국 자생종인 속리기린초(P. zokuriensis)와 태백기린초(P. latiovalifolium)의 완전한 엽록체 유전체를 Illumina 시퀀싱 기술을 이용하여 해독하였다. 분석 결과, 두 종은 유사한 유전체 크기와 구조를 보였으며 각각 88개의 단백질 코딩 유전자, rRNA 및 tRNA 유전자를 포함하였다. 그러나 일부 tRNA 유전자의 차이가 확인되어 종 구분을 위한 잠재적 분자 표지로 활용될 수 있음을 시사하였다. 계통학적 분석에서는 두 종이 모두 Phedimus 속에 속하면서 Sedum 속과 뚜렷이 분리되어 독립된 진화 계통임이 확인되었다. 또한 비교 분석을 통해 총 57개의 InDel과 168개의 SNP 변이지역이 확인되어 향후 종 동정, 집단유전학 및 보전 연구를 위한 기초 자료로 활용 가능성이 제시되었다. 본 연구는 한국 자생 기린초속 식물의 분류학적 위치와 유전적 다양성 이해를 심화시키고, 이들의 보전 및 경제적 활용을 위한 중요한 자원을 제공한다.
