

Introduction
Materials and Methods
Gymnosperm species, permanent voucher slides housed, and anatomical terminology
Cutting & softening wood block, microtome section and observation
Results
Anatomical characteristics required for the identification of some Korean gymnosperm sawn timber
An identification key of some Korean gymnosperm sawn timber based on anatomical characters
Discussion
Introduction
Most of the forest trees in Korea are mature, reaching 30 to 50 years of age (Korea Forest Service, 2021), so their growth rate gradually slowing and carbon fixation decreasing (Greenhouse Gas Information Center, 2020). In forest, trees account for the largest proportion of biomass at 82%, and 50% of which is carbon dioxide accumulated in their cell walls (Arsić et al., 2021). It now becomes urgent not only to process and utilize felled trees for long-term carbon storage, but also to improve forest trees as more valuable species (Kim et al., 2023; Korea Forest Service, 2021, 2024).
There are not many anatomical reference materials as well as trained students on the Korean gymnosperm sawn timber, so it is not going to easy in identifying them when available only sawn timber or wooden goods. Lee and Eom (1984, 1987) had reported two articles about the “Wood identification of the veneer species that grow in Korea”. These two reports were evidenced by characteristics and identification by gross features by naked eyes and microscopic features respectively. Lee and Chun (1990) reported a study on identifying and searching Korean woods using a computer program. However, to improve the accuracy of this computer program, anatomical database covering more species are needed. Additionally, several articles have used anatomical and other tools to identify a limited number of trees species (Chu, 2024; Kim et al., 1992; Kim and Park, 1994; Lee et al., 2009; Park et al., 1994; Yang et al., 2012). The book named “Illustrated Conifers of Korea III” (Yang et al., 2012) included almost all of plant characteristics about cone, seed, pollen, leaf, life cycle, simple anatomy, systematic analysis etc. However, if there are available only sawn timber without the generative and vegetative features mentioned above, the comprehensive anatomical data is necessary to identify gymnosperm species.
In addition, there are several studies that identify the species and their uses of various wooden objects found from both various archaeological sites in Korea (Hwang et al., 2020; Park, 1922, 1993; Park et al., 2022). The wood silicification has been viewed abundance in fossil record, together with a long evolutionary history of woody plants (Mustoe, 2023). Kim et al. (2002) investigated the anatomical characteristics of several siliconized conifer woods and attempted to restore the paleovegetation and paleoclimate from Korea of the Cretaceous period. However, the reference materials of anatomical features much more needed in order to identify both the various wooden household items found in archaeological sites, and also the reconstruct of paleoenvironment using silicified woods found in geological sites.
The study aims to figure out the anatomical characteristics on some Korean gymnosperm, and then to create an identification key for species of the sawn timber. The above data might also be used as reference material when identifying wooden household items discovered in archaeological and paleological study.
Materials and Methods
Gymnosperm species, permanent voucher slides housed, and anatomical terminology
The materials used in this study were 18 species of gymnosperms (belonging to seven families and 18 genera; Park, 2007) (Table 1). Their wooden materials and permanent voucher slides are housed in the Botany Laboratory, Division of Science Education, Jeonbuk National University, Korea. The anatomical characteristics and their terminology of the wood basically followed the guidelines of ‘International Association of Wood Anatomists’ (IAWA, 2004; Park et al., 1981), and a bit were also changed in this study when necessary.
Table 1.
List of gymnosperm species and their permanent voucher slides used in this study.
Cutting & softening wood block, microtome section and observation
The collected woods were cut into 1 cm3 cube blocks able to see three-dimensional sides, and embedded in polyethylene glycol (PEG2000). Some of the wood blocks were softened by boiling in a pressure cooker for different times depending on their strength. And then, the softened wood blocks were preserved in the 1:1:1 (v/v) mixture of absolute ethanol, glycerin, and water until sectioning. Using a sliding microtome (Thermo Shandon Limited, UK), they were cut into transverse, tangential, and radial views with a thickness of 10-15 ㎛. The cut sections were immediately transferred to 50% alcohol, and then stained in a staining jar with fast green for 20 seconds and safranine for more than 2 hours respectively (Bond et al., 1980; Japan Wood Research Society, 1985). One section of each of the three directions was put on a slide glass, on that 2-3 drops of permount medium were treated, and the slides were finally sealed with a cover glass to make a permanent preparation. The images of anatomical characteristics were observed, measured, and photographed using an optical microscope (AxioLab microscope, Carl Zeiss).
Results
Anatomical characteristics required for the identification of some Korean gymnosperm sawn timber
To make an identification key on some Korean gymnosperm sawn timbers, anatomical features were investigated in the level of cell and tissue by a compound microscopy. The seven major characters were firstly selected: intercellular canals (or resin ducts), axial parenchyma cells, axial tracheid, ray tracheid, ray parenchyma cells, cross-field pitting, and ray tissue (Table 2). These major characters were further divided into few detailed characters. The character states of each detailed characters were finally described together with micrographs (Figs 1, 2, 3, 4, 5, 6).
Table 2.
Characteristics in creating an anatomical key of some Korean useful gymnosperm woods.
| Characterz | Detailed characterz | Character statesz |
|
Axial intercellular Resin duct (or canals) in transverse view | Present or absent | Present/absent |
| Thickness of epithelial of intercellular resin duct | Thick-walled/thin-walled | |
|
Axial parenchyma cell in tangential view | Present or absent | Present/absent |
| Arrangement | Diffuse/tangentially zonate | |
| Transverse end walls | Smooth/beaded or nodular | |
|
Axial tracheid in tangential view | Helical thickening in tracheid | Present/absent |
|
Spacing (more or less than 120/㎜) and shape of helical thickening |
Single/double or triple/narrowly spaced/widely spaced | |
| Tracheid pitting in radial walls | Uniseriate/two or more seriate | |
|
Ray tracheid in radial view | Present or absent | Present/absent |
| Cell wall shape | Smooth/dentate | |
|
Ray parenchyma cell in radial view | Moniliform at end walls | Present/absent |
| Indentures | Present/absent | |
|
Cross-field pitting in radial view | Shape | Fenestriform/pinoid/piceoid/ Cupressoid/taxodioid |
| No. of pits per cross-field | No. Of 1-2/1-3/3-5 | |
|
Ray parenchyma in tangential view | Ray width by cells | Exclusively uniseriate/2-3 seriate in part |
| Average ray height by no. Of cells | Very low (>4 cells)/medium (5-15)/high (16-30) | |
| Fusiform ray parenchyma | Present/absent |
The axial intercellular canals in the transverse view (from here on, TrV) include two states; present (Figs 1A, B) or not, and the thin (Fig. 1A) or thick (Fig. 1B) of the thickness of the epithelial cell walls. The axial parenchyma cells in TrV have two states; present (Figs. 1C, D) or absent, and a diffused (Fig. 1C) or tangentially zonate arrangement (Fig. 1D). In the tangential view (from here on, TaV), the transverse walls of axial parenchyma have two states; smooth (Fig. 2A) or beaded types (Fig. 2B). In TaV, the spiral thickening of tracheid walls has two states; a single (Fig. 2C) and double rows (Fig. 2D). In the radial view (from here on, RaV), the axial tracheid has two states of bordered pits; a single (Fig. 3A), and single and/or double arrangement (Fig. 3B). In RaV, the radial tracheid has two states; present (Figs. 3C, D) or absent of beaded end walls (Fig. 3C). In RaV, the lateral walls of radial tracheid have three states; smooth (Fig. 3D), dentate (Fig. 3E), and reticulate thickening lateral walls (Fig. 3F). In RaV, the moniliform end walls of ray parenchyma cells have two states; smooth (Fig. 4A) and beaded type (Fig. 4B). In RaV, the shape of cross-field pitting developed on parenchyma cell walls has five states (Fig. 5F); fenestriform (Fig. 5A), pinoid (Fig. 5B), piceoid (Fig. 5C), cupressoid (Fig. 5D), and taxodioid (Fig. 5E).

Fig. 1.
Micrographs of resin duct (A, B) and axial parenchyma arrangements (C, D) type in cross view. A. Epidellial cells with thin-walled in Pinus sylvestris (arrow). B. Epidellial cells with thick-walled in Larix kaempferi (arrow). C. Diffuse in Cephalotaxus koreana (arrows; blue circular cells). D. Tangentially zonate to the annual ring direction in Platycladus orientalis (arrows; dark red cells).
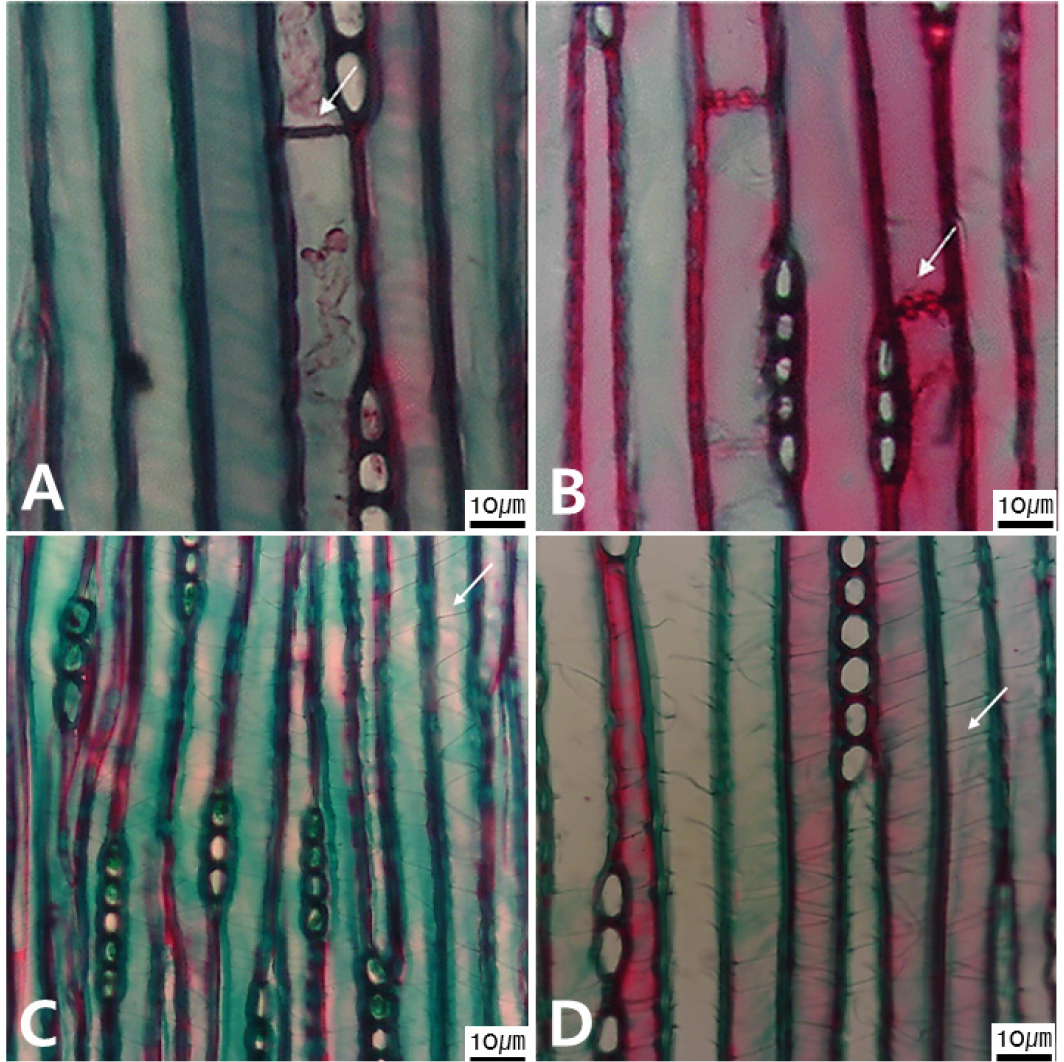
Fig. 2.
Micrographs of both the transverse end walls of the axial parenchyma cell (A, B) and the spiral thickening of the tracheid walls (C, D) types observed in tangential view. A. Smooth (arrow) in Cunninghamia lanceolata. B. Beaded in Taxodium distichum (arrow). C. A single row in Taxus cuspidate (arrow). D. Two rows in Torreya nucifera (arrow).

Fig. 3.
Micrographs in radial view showing of both pitted arrangement of axial tracheid (A, B), and terminal wall & lateral walls (C-F) types of radial tracheid. A. Single row in Picea jezoensis (arrow). B. 1-2 rows in Metasequoia glyptostroboides (arrows) C. Beaded end in Larix kaempferi (arrow). D. Smooth in Pinus strobus (arrow). E. Dentate in Pinus rigida (arrow). F. Reticular thickening in Pinus banksiana (arrow).


In TaV, the radial parenchyma shows three characters; ray width by cells (Figs. 6A, B), average ray height by the number of cells (Figs. 6D-F), and fusiform ray parenchyma (Fig. 6C). The ray width by cells has two states; exclusively uniseriate (Fig. 6A) and 2-3 seriate type (Fig. 6B). The average ray height by the number of cells shows three states; very low below w/ four (Fig. 6D), medium w/ five to fifteen (Fig. 6E), and high w/ sixteen to thirty cells (Fig. 6F). The fusiform ray parenchyma has two states; present (Fig. 6C) or absent.
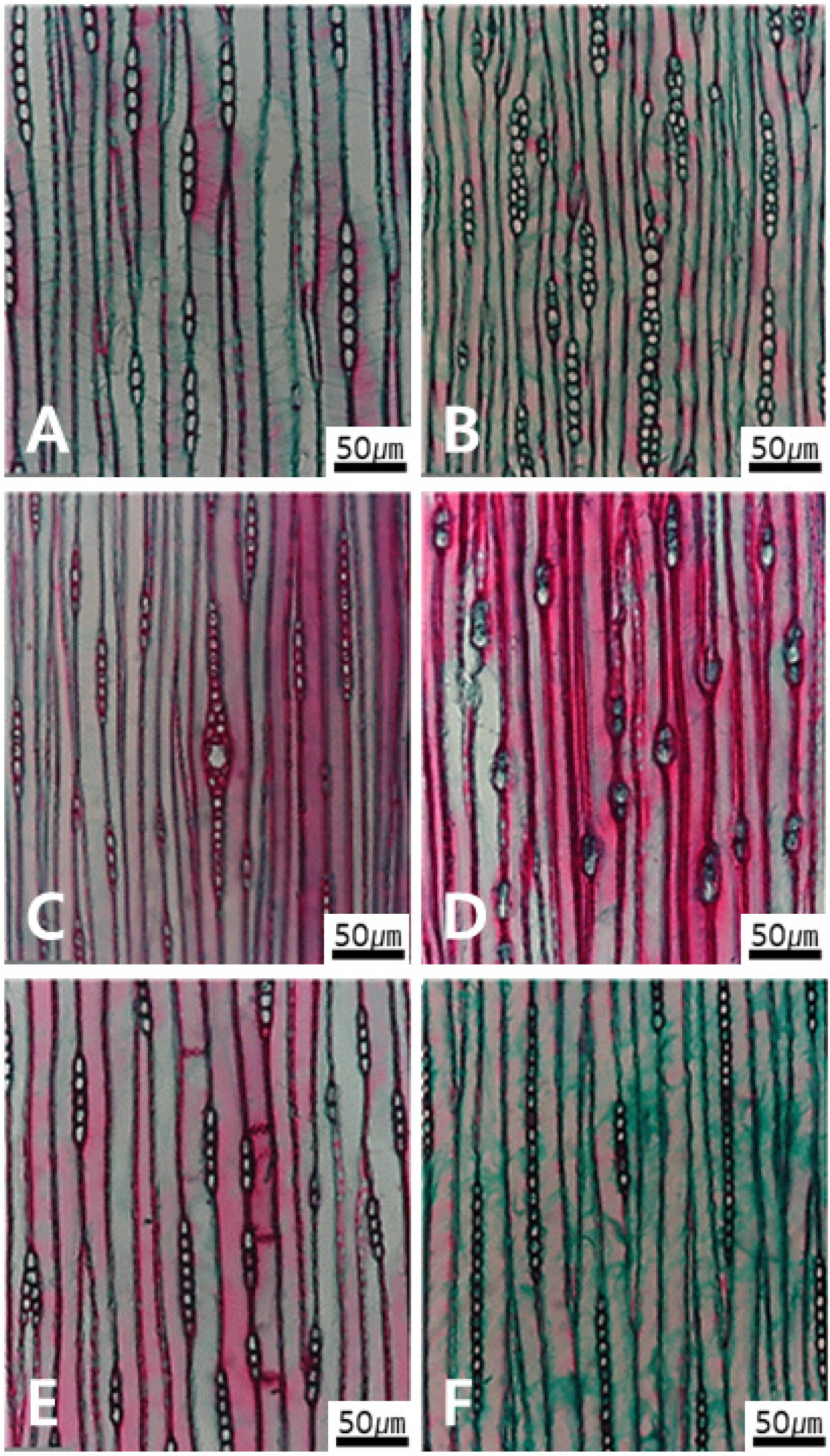
Fig. 6.
Micrographs of radial parenchyma cells in tangential view. A. Ray cell width showing single-row in Torreya nucifera. B. 1-2 rows in Chamaecyparis obtusa. C. Present of fusiform ray parenchyma in Picea jezoensis. D-F. Average ray height by number of cells having three types. D. Very low with 1-4 cells in Ginkgo biloba. E. Medium with 5-15 cells in Taxodium distichum. F. High with 16-30 cells in Abies holophylla.
An identification key of some Korean gymnosperm sawn timber based on anatomical characters
This identification key was created for the purpose of species identification of sawn timber, rather than for botanical taxonomic purposes. Therefore, it is not insisting on a binary classification system, but created for accurate and convenient use when identifying the Korean gymnosperm wood.
1A. Present of ray tracheid in radial view 2
2A. Smooth ray tracheid walls Tsuga sieboldii
2B. Dentate ray tracheid walls 3
3A. Uniseriate pitting only in radial walls 4
3B. Mixed of uni- and di-seriate pitting of in radial walls Larix kaempferi
4A. Pinoid cross-field pitting between axial tracheid and ray Pinus densiflora
4B. Piceoid cross-field pitting between axial tracheid and ray Picea jezoensis
1B. Absent of ray tracheid in radial view 5
5A. Present of axial parenchyma cell in transverse view 6
6A. Diffused arrangement of axial parenchyma cell 7
7A. Uniseriate axial-tracheid pitting 8
8A. Ray parenchyma with 1~4 cell height in tangential view Thujopsis dolabrata
8B. Ray parenchyma w/ 5~15 cell height in tangential view Cunninghamia lanceolata
7B. uni- or di-seriate axial-trachied pitting 9
9A. Ray parenchyma with 1~4 cell height in tangential view Ginkgo biloba
9B. Ray parenchyma w/ 5~15 cell height in tangential view Metasequoia glyptostroboides
6B. Coexist of diffused & tangentially zonate of axial parenchyma 10
10A. Axial parenchyma end walls w/ smooth Cryptomeria japonica
10B. Axial parenchyma end walls w/ beaded or nodular Taxodium distichum
10C. Axial parenchyma end walls w/ coexist of smooth and beaded Cephalotaxus koreana
6C. Tangentially zonate arrangement of axial parenchyma cell 11
11A. Axial parenchyma end walls w/ smooth Chamaecyparis obtusa
11B. Axial parenchyma end walls w/ beaded or nodular Juniperus chinensis
11C. Axial parenchyma end walls w/ coexist of smooth and beaded Platycladus orientalis
5B. Absent of axial parenchyma cell in transverse view 12
12A. Absent of axial tracheid w/ helical thickening in radial view 13
13A. Axial tracheid w/ single helical thickening Taxus scuspidata
13B. Axial tracheid w/ double to triple helical thickening Torreya nucifera
12B. Present of axial tracheid w/ helical thickening in radial view 14
14A. Fenestriform cross-field pitting between tracheid and ray Sciadopitys verticillata
14B. Taxodioid cross-field pitting between axial tracheid and ray Abies holophylla
Discussion
The anatomical characters considered as a pivotal for species identification could little bit be different from each other depending on the researchers (IAWA, 2004; Pandey, 2021; Park et al., 1981). This study had done by two main steps. we tried at first to figure out the efficient anatomical characters in identifying the species of sawn timber, and then create an identification key mainly using their characters. The characters selected for the creation of the identification key were as follows: presence/or not and arrangement of axial parenchymal cells, presence/or not and the shape of end walls of axial parenchyma cell, arrangement of axial tracheid pitting, presence/or not and the end walls shape of ray tracheid, cross-field pitting in radial view, height of ray parenchyma in tangential view (Table 2).
The identification key developed here has two major features. First, this study described each of a single directional view to figure out exactly characters and its states as to every cell and tissue features. This is because the shape of plant cells often has a very different appearance depending on the viewing direction such as transverse, radical, and tangential section. For example, the arrangement of the axial parenchyma cells in the gymnosperm wood is easily observed in transverse view (Fig. 1C, D), but the end walls shape of the same tissue and cell is more easily observed in tangential view (Fig. 2A, B). Even within the same tissue or cell, different directional sections are suitable for observing anatomical characteristics, so clearly indicating these features facilitates wood identification. Second, the key was created by selecting features that are easier to investigate. While the presence/or not of indentations in Sciadopitys verticillata and Abies holophylla can be distinguished between species (IAWA, 2004; Park et al., 1981; Yang et al., 2012), but the easier-to-investigate shape of the cross-field pitting was used to distinguish them (Fig. 4B, Fig. 5). Rather than focusing on difficult tissues or features that can be easily confused by beginners, we primarily utilized features that can be clearly observed characteristics under a microscope.
In general, Pinaceae spp., belonging to the genera Larix, Picea, Pinus, and Tsuga etc, produce radial tracheid (IAWA, 2004). Their morphology of the cell walls forming radial tracheid has shown to be an important characteristic for their identification and creating anatomical key (Fig. 3A, D–F). Unlike angiosperm wood, gymnosperm exhibit radial cross-field pitting, formed by the intersection of radial parenchyma cells and axial tracheid in a radial cross-section (IAWA, 2004). The cross-field pitting patterns here appeared five anatomical character states, which served as an important feature for identification of gymnosperm wood (Fig. 5). The five are: Fenestriform type in Sciadopitys verticillate and Pinus densiflora, Piceoid in Picea jezoensis, Taxodioid in Abies holophylla, and Cupressoid type in Torreya nucifera (Fig. 5, Table 2).
In addition, this study conducted because of still the lack of comprehensive reference materials for identifying gymnosperm timbers in Korea. Several previously reported anatomical studies on gymnosperm wood are assumed that specific aims using limited species rather than detailed descriptions of the anatomical features of tissues and cells by attaching microphotographs (Lee and Eom, 1984, 1987; Park et al., 1981). Park (1992) had described four gymnosperm species from the archeological site. But he did not describe their anatomical characteristics of species, but only describe ecological features at that time. Hwang et al. (2020) had reported the wood fragments of four species from the wooden members of the Manseru Pavilion at Bongjeongsa Temple in Andong of Korea. They were adopted Synchroton X-ray Microtomography-Based Three-Dimensional Microstructural Imaging to identify the objects with highly limited sampling capabilities such as important and registered wooden cultural properties.
In particular, Pandey (2021) had reported an article named “Climatic influence on tree wood anatomy: a review”. He investigated on the influence of climatic parameters on wood anatomy from the different geographical regions using recent (2015~2020) published articles on the topic. He mentioned the relationship between the anatomical features of gymnosperm and angiosperm xylem with climatic factors. And then he proposed to need more study for the temporal and spatial variation in the anatomical characteristics of xylem tree rings with environmental changes.
Meanwhile, several multidisciplinary studies related to the wood features have been conducted recently. MacLeod (2025) recently wrote a book named “Archaeological Wood and Woodworking”. He introduced the study of archeological wood relating to wood materials, wood anatomy, and dendrochronology etc. Pandey (2021) reviewed the relationship between the anatomical features of gymnosperm xylem with climatic factors using the literatures published during 2015~ 2020. He in particular reported the implications of spatiotemporal variations in the anatomical characteristics of xylem annual rings. When identifying plant macro-fossils, plant phylogenetic research was also conducted by combining anatomical characteristics and other data (Kim et al., 2002). For example, Xie et al. (2025) had investigated the Mesozoic gymnosperm Xenoxylon Gothan, and they had reported that the fossil reveals close affinity with extant Podocarpaceae (Coniferales). They had combined the data sets of genes, xylological and biomolecular character to reach above results. The wood record of archeological sites is a ubiquitous element. It is also true that the anatomical reference is necessary for identifying the wood record.
