

서 언
뽕나무과(Moraceae) 닥나무속(Broussonetia)의 수종들은 전통적 종이 제조의 중요한 원료로 사용되어 왔다(Kim et al., 2019). 닥나무속(Broussonetia)은 동남아시아와 동아시아에 걸쳐서 5종이 분포하고 있는데, 우리나라에는 애기닥나무(Broussonetia kazinoki Siebold ex Siebold & Zucc)와 꾸지나무(B. papyrifera (L.) L’ Hér. Ex Vent.), 닥나무(B. × kazinoki Siebold) 등 3종이 자생하고 하고 있다(Chung et al., 2017). 애기닥나무는 우리나라를 비롯하여 중국과 대만, 일본에만 분포하는 동아시아 지역 특산식물로서, 자연 교잡종인 닥나무(B. × kazinoki)의 양친중 모계(maternal parent)이다(Kim et al., 2019). 자웅이주(Dioecy)인 꾸지나무와 닥나무와는 달리 자웅동주(Monoecy)로서 4월 중순~5월 초순에 개화하고, 6월말~7월 초순에 결실한다. 열매는 집합과로서 둥글고 지름 1~1.5 ㎝ 크기로 붉게 익으며, 새들에 의해 주로 산포되고, 하나의 열매에 1.3~1.5 ㎜ 크기의 10~30개의 종자가 들어 있다.
최근에는 전세계 제약회사들이 천연물을 이용하여 신약개발에 중요한 원천소재로써 사용하고 있으며, 야생식물의 유용 생리활성에 관한 연구를 통해 신약개발 산업화가 증가되는 추세이다(Yoo et al., 2016). 애기닥나무 줄기에서 추출한 B. kazinoki stem extract (BKSE)를 통해 collagenase 활성을 억제하여 피부의 콜라겐 함량을 유지하는데 도움을 줄 수 있다고 보고되었다(Kwon et al., 2019). 또한 열매에서 추출한 B. kazinoki fruit extract (BKFE)는 당뇨병을 예방할 수 있다고 알려져 있으며(Kim et al., 2020), 잎은 아토피 피부염을 완화하는데 효과가 있다고 알려져 있기 때문에(Lee et al., 2014), 부위별로 이용 가치가 높다는 것을 알 수 있다. 식물 소재를 산업화하기 위해서는 대량 증식 방법이 확립되어야 하는데(Shin et al., 2017), 이를 위해 번식 프로토콜을 만드는 것이 중요하다.
휴면을 가진 종자는 발아하기에 유리한 환경이 조성되기 전까지 지연시간을 가진다(Graeber et al., 2017). 야생의 종자는 다양한 휴면유형을 가지고 있으며, 종자휴면의 종류로는 생리적 휴면(physiological dormancy, PD), 형태적 휴면(morphological dormancy, MD), 형태생리적 휴면(morphophysiological dormancy, MPD), 물리적 휴면(physical dormancy, PY), 조합휴면(combinational dormancy, PY+PD)으로 크게 다섯 가지로 나뉘어져 있다(Baskin and Baskin, 2004). 이러한 종자의 휴면을 타파하기 위해서는 고온층적, 저온층적, 식물생장조절물질 등의 인위적인 처리를 해야 한다(Baskin and Baskin, 2004; Bewley and Black, 1982). 이외에도 휴면(dormancy)과 비휴면(non-dormancy)의 전환 단계로서 종자가 좁은 범위의 발아 적온에서만 발아하는 conditional dormancy(CD) 유형이 있다(Hartmann et al., 2002).
애기닥나무는 뽕나무과(Moraceae)에 속하며, 뽕나무과(Moraceae)의 종자 내부 형태는 배가 구부려져 있으며, 완전히 발달된 배를 가지고 있다고 보고되었다(Barbour et al., 2008). 닥나무속(Broussonetia)에 속하는 중국의 Broussonetia papyrifera 종자의 발아 최적온도는 30℃로 보고되었다(Yan et al., 2019). 터키에서는 근연종인 Morus nigra L. 종자를 연구하여, 대조구에서는 33%의 발아율을 보인 반면, 100일 동안 저온층적처리를 한 결과 88%의 높은 발아율을 나타냈다고 보고하였다. 또한 GA3처리를 통해, 1000 ㎎/L에서 60%의 발아율을 보이며 휴면이 타파되었다(Koyuncu, 2005). 미국에서는 Morus rubra 종자를 발아시키기 위해서는 광이 요구되며(Dirr and Heuser, 1987), 30일 동안의 저온층적처리를 통해 88%의 높은 발아율을 보였다고 하였다(USDA FS, 2002). 그러나 애기닥나무의 종자번식에 대한 연구는 없었다.
자생식물에 대한 번식 프로토콜 데이터베이스를 북미에서 구축하고 있으며(Dumroese and Landis, 2016), 이를 통해 대량 증식 방법을 확립하고 있다. 국내의 경우도, 자생식물의 종자기반 대량번식 프로토콜을 만들기 위해서 지속적인 기초자료의 확보가 필요하다. 본 연구는 기능성 소재 자원으로 유망한 애기닥나무 종자의 휴면과 발아특성을 조사하고, 추후 대량번식에 활용할 수 있는 기초자료를 제공하고자 수행하였다.
재료 및 방법
식물 재료
실험에 사용된 애기닥나무 종자는 2019년 6월 15일 국립수목원에서 수집되었다(Table 1, Fig. 1). 수집한 후 실험실 내(21~25℃)에서 2019년 7월 6일까지 건조하였다. 건조된 종자는 실험에 사용되기 전까지 저온저장고(mean ± SE: 0 ± 0.5℃)에 보관하였다.
Table 1.
Seed characteristics of Broussonetia kazinoki Siebold ex Siebold & Zucc

기본 특성
버니어캘리퍼스를 이용하여 종자의 길이와 너비를 측정하였다. 종자의 무게는 100립 중(㎎)으로 측정하였다. 종자의 내·외부 형태 조사를 위해 종자를 면도칼을 이용하여 종단으로 자른 후 USB현미경(AM3111 Dino-Lite premier, ANMO Electronics Co., Taiwan)을 사용하여 관찰하였다. 종피에 불투수성이 있는지 판단하기 위하여 수분흡수 실험이 진행되었으며, 90 × 15 ㎜의 Petridish에 여과지 2매를 깔고 증류수를 충분히 주입한 후 20립 3반복으로 치상하였다. 치상된 종자는 0, 3, 6, 9, 12, 24, 시간 후의 무게가 조사되어 수분흡수율이 계산되었다. 수분흡수율은 %Ws = [(Wh-Wi)/Wi] × 100을 통해 계산되었으며(Lee et al., 2015), Ws = 증가된 종자 무게의 비율, Wh = 수분이 흡수된 종자의 무게, Wi = 초기의 종자 무게를 의미한다.
광·온도 조건
애기닥나무 종자를 소독하기 위해 500 ㎎/L 베노밀(Benomyl, FarmHannong, Seoul, Korea)에 24시간 침지 처리한 후 Petridish (90 × 15 ㎜)에 여과지 2매를 깔고 증류수를 충분히 주입한 후 20립 4반복으로 치상하였다. 모든 Petridish는 수분손실 방지를 위해 파라필름으로 밀봉하였다. 광조건은 Petridish를 12/12 h(광조건/암조건)의 환경에 두었으며, 광조건 하에 조사한 광의 PPFD는 63.46 μmol·m-2·s-1이었다. 암조건은 광조건과 동일한 환경이지만 Petridish를 알루미늄 호일로 감싸 암조건을 형성하였다. 온도 조건은 4℃로 설정된 콜드랩 챔버(HB-603CM, HANBAEK-Scientific, Korea, Bucheon)에, 15/6, 20/10, 25/15℃는 멀티룸 챔버(HB-302S-4, HANBAEK-Scientific, Korea, Bucheon)에 완전임의 배치하여 배양하였다. 배양한 종자는 6주 동안 발아율을 조사하였으며, 발아는 유근이 1 ㎜ 이상 돌출하였을 때를 기준으로 하였다.
GA3 처리
종자를 실온(22~25℃)에서 GA3용액 0(증류수), 10, 100, 1,000 ㎎/L의 농도에 24시간 침지 후 5시간 동안 베노밀 500 ㎎/L 용액에 소독하였다. 소독된 종자들은 Petridish (90 × 15 ㎜)에 여과지 2매를 깔고 증류수로 충분히 주입한 후 20립 4반복으로 치상하였다. 치상된 Petridish는 15/6, 25/15℃ 멀티룸 챔버에 완전임의 배치하여 배양하였다. 배양한 종자는 6주동안 발아율을 조사하였으며, 발아는 유근이 1 ㎜ 이상 돌출하였을 때를 기준으로 하였다.
저온층적처리
저온층적처리의 휴면타파 및 발아촉진 효과를 알아보기 위해 광·온도 조건 실험과 동일한 방법으로 소독한 다음 Petridish (90 × 15 ㎜)에 여과지 2매를 깔고 증류수로 충분히 주입한 후 20립 4반복으로 치상하여 0, 4, 8, 12주 동안 저온층적을 실시하였다. 저온층적처리에는 4℃로 설정된 콜드랩챔버에 완전임의 배치하여 배양하였다. 저온층적처리가 끝나면 25/15℃의 변온으로 이동하여 배양하였다. 배양한 종자는 4주 동안 발아율을 조사하였으며, 발아는 유근이 1 ㎜ 이상 돌출하였을 때를 기준으로 하였다.
통계 처리
각 발아시험 결과에 대하여 조사된 데이터는 SAS 9.4 (SAS Inst. Inc., Cary, NC, USA)를 이용하여 분산분석을 하였고, 처리 평균간 비교는 Tukey’s HSD test (p < 0.05)로 검정하였다. 그래프는 SigmaPlot 10.0 (SPSS Inc., Chicago, IL, USA)이 사용되었다.
결 과
기본 특성
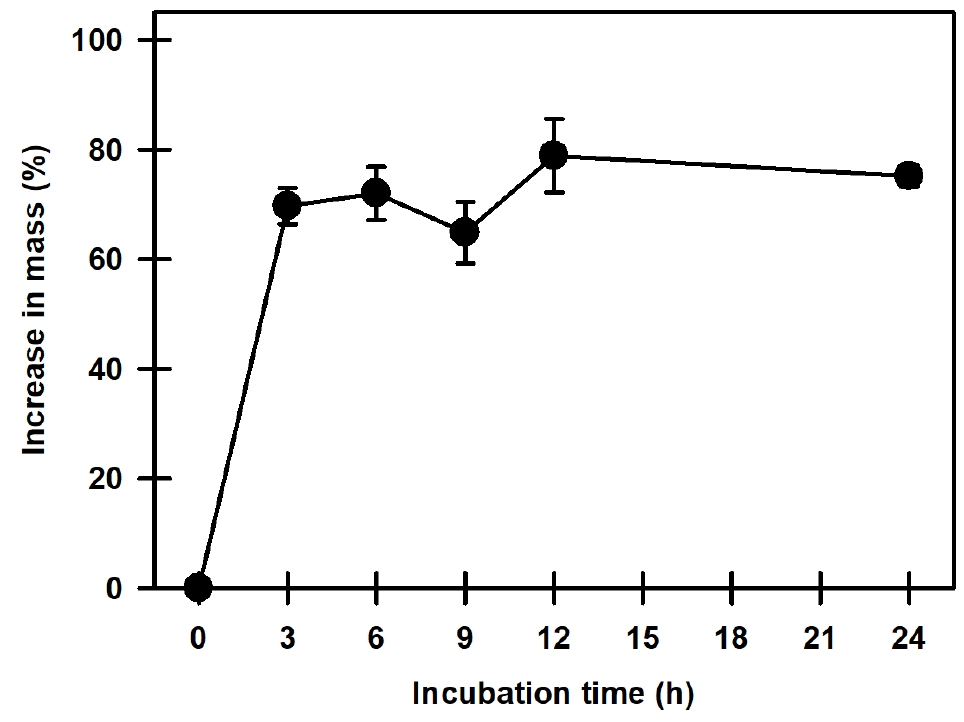
애기닥나무의 종자 길이는 1.44 ± 0.01 ㎜, 너비는 1.20 ± 0.01 ㎜, 100립중은 71.68 ± 0.83 ㎎이었다(Table 1). 종자의 외부형태는 타원형의 옅은 노란색을 띠고 내부 형태는 구부려진 굽은(bent) 형태로, 발달된 배를 가지고 있었다(Fig. 2). 수분흡수 실험에서는 종자의 무게가 3시간만에 초기 종자 대비 60% 이상 증가하였다(Fig. 3).

Fig. 2.
Process of germination and seedling development of Broussonetia kazinoki. Bars = 1 ㎜. (A) seed outside morphology, (B) a longitudinal section of a seed with fully-developed embryo, (C) an emerged radicle; (D-E) subsequent developmental stages, (F) cotyledons morphology, (G) a seedling. ec, endocarp; en, endosperm; sc, seed coat; em, embryo; co, cotyledons; ep, epicotyl.
광·온도 조건의 영향
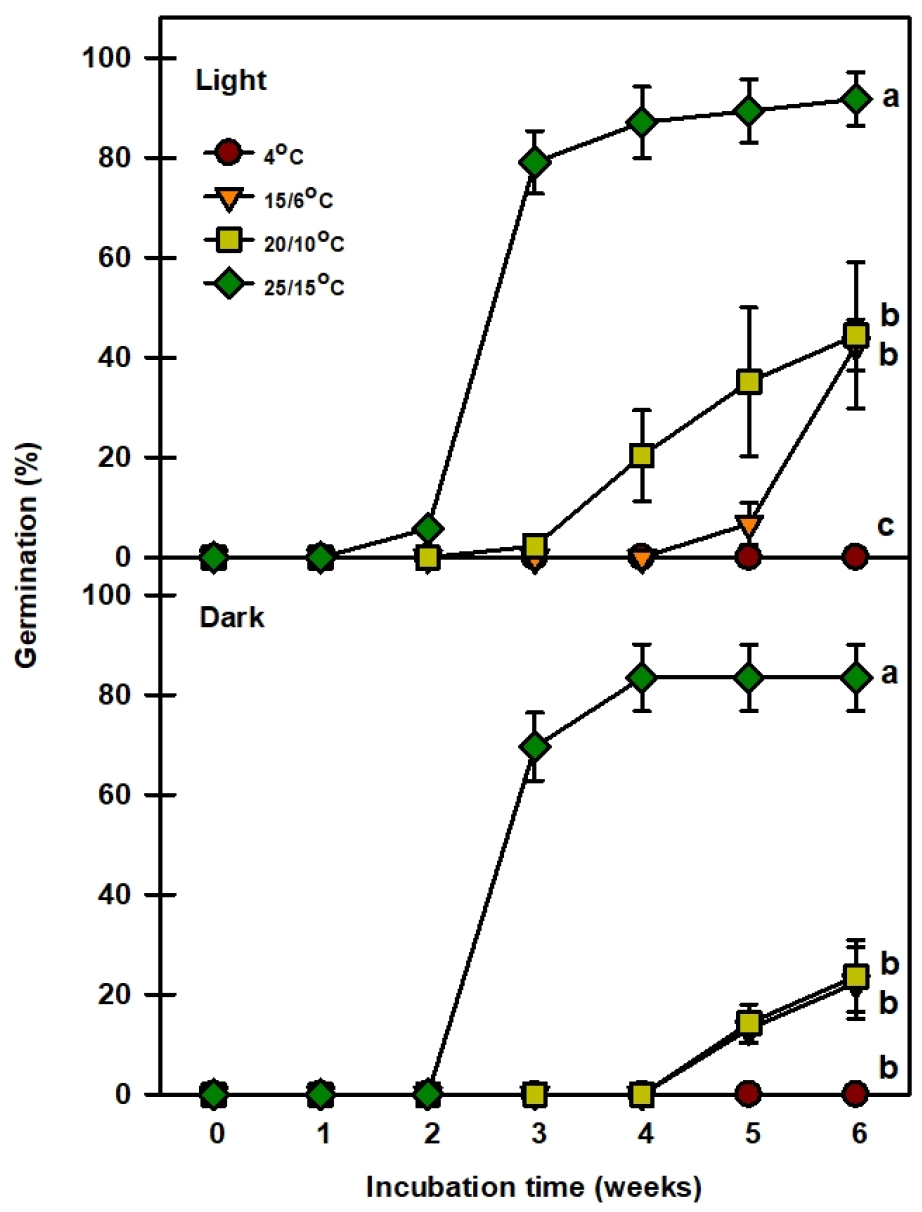
명조건 4℃에서는 발아를 하지 않고, 15/6℃에서 5주차에 발아가 시작하여 최종 발아율이 42.5%를 보였다(Fig. 4). 20/10℃의 경우 3주차에 발아가 시작하여 최종 발아율이 44.4%로 15/6℃와 최종 발아율은 비슷했지만 발아속도가 빠른 것을 알 수 있었다. 25/15℃에서는 2주만에 발아가 시작되어 최종 발아율이 91.7%로 가장 발아율이 높은 것을 알 수 있으며, 이를 통해 명조건에서는 25/15℃가 최적온도임을 알 수 있었다. 암조건에서는 명조건과 마찬가지로 4℃에서는 발아를 하지 않고, 15/6, 20/10℃에서 5주차에 발아를 시작하여 최종 발아율이 각각 22.3, 23.6%를 보이며 발아율이 낮은 것을 알 수 있었다. 하지만 25/15℃에서는 3주만에 발아를 시작하여 최종 발아율이 83.4%로 암조건에서 가장 발아율이 높았다. 본 연구결과 온도에 따른 발아율의 차이가 있음을 알 수 있고, 광조건에 따른 차이는 4℃와 25/15℃를 제외하고 다소 차이가 있음을 알 수 있었다.
GA3 처리의 효과
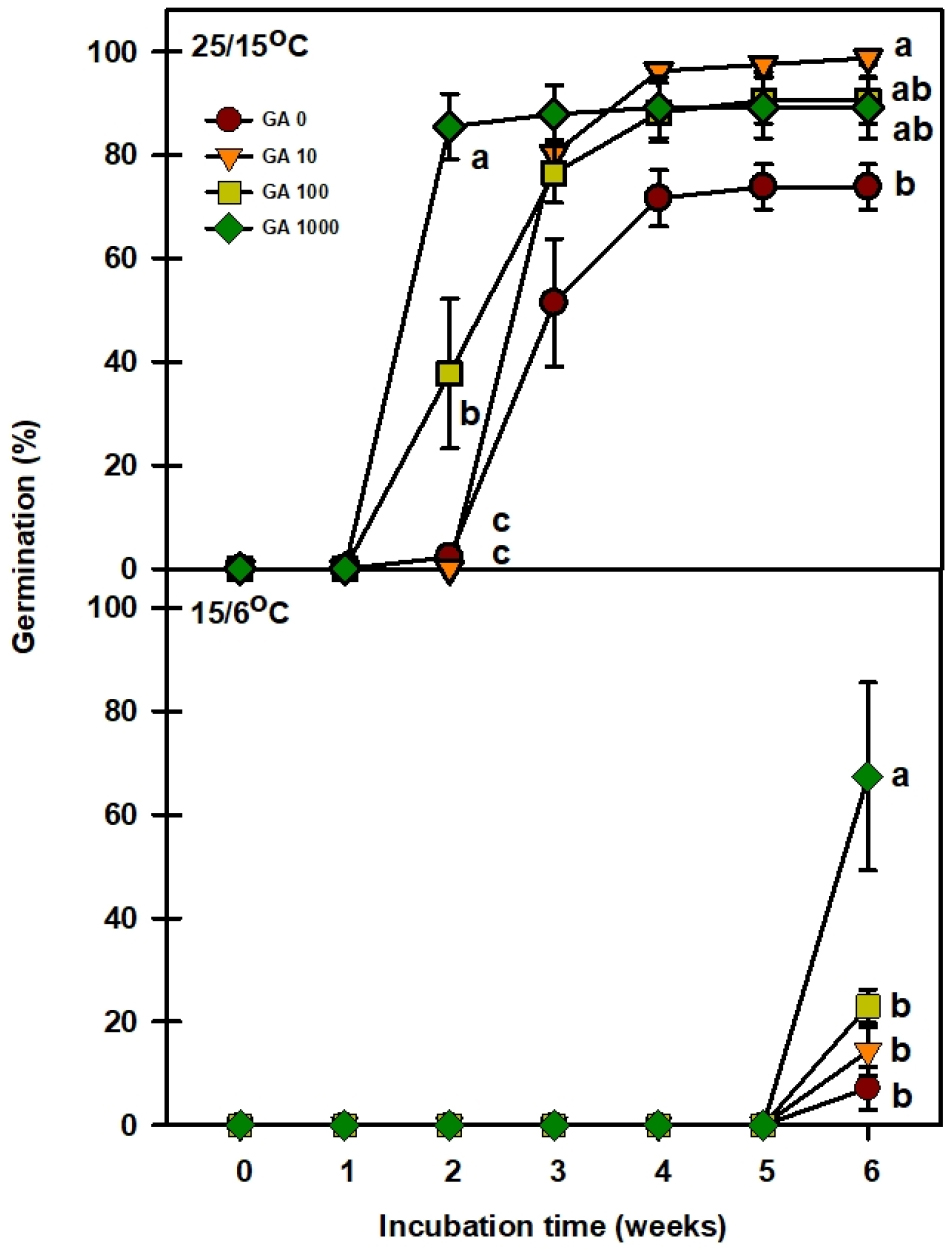
농도별 GA3를 처리한 결과 15/6℃에서 모든 처리구에서 5주차까지는 발아를 하지 않고 6주차에 발아한 것을 보였다(Fig. 5). 0, 10, 100 ㎎/L에서는 최종 발아율이 7.1, 14.2, 22.9%을 보이며, 농도가 높을수록 발아율이 높아지는 것을 알 수 있었다. 1000 ㎎/L 농도처리에서는 최종 발아율이 67.3%로 15/6℃에서 가장 높은 발아율을 보였다. 25/15℃에서는 대조구가 3주차에 발아를 시작하여 73.8%의 최종 발아율을 보였다. 10 ㎎/L 농도처리에서는 3주차에 발아를 시작하여 98.7%의 최종 발아율을 보였으며, 100, 1000 ㎎/L 농도처리에서는 2주차에 발아를 시작하여 각각 90.5, 89.1%의 최종 발아율을 보였다. 본 연구결과 농도처리가 높을수록 발아소요 일수를 앞당기는 효과를 알 수 있었다.
저온층적처리의 효과
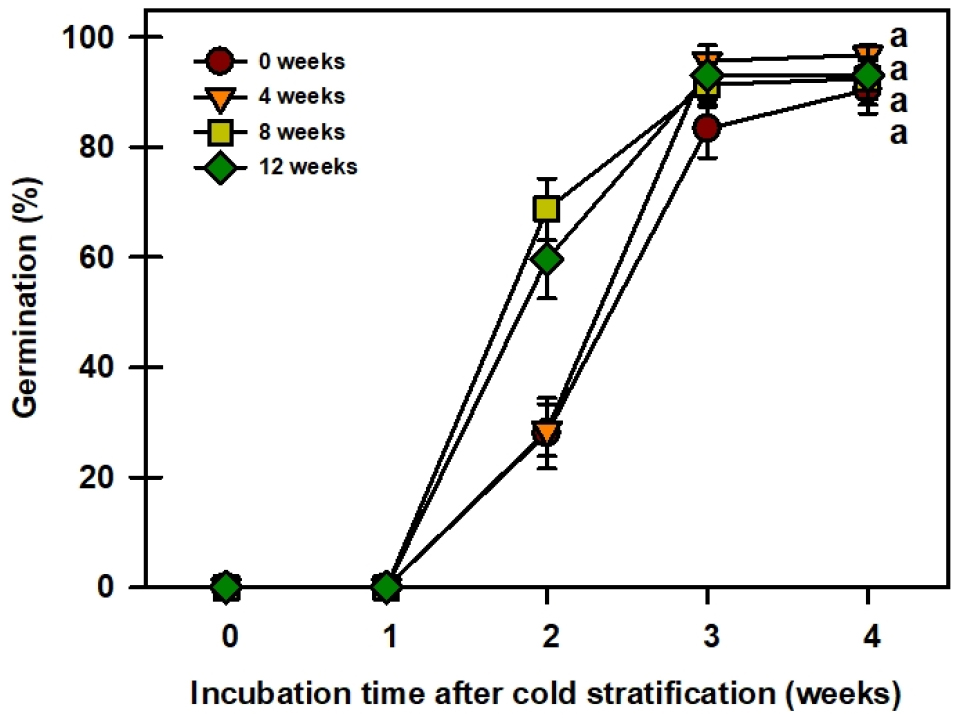
저온처리 기간별 실험결과 모든 처리구에서 2주차에서 발아를 시작한 것을 알 수 있다(Fig. 6). 대조구와 저온 4주처리는 2주차에 발아율이 각각 27.9, 28.5로 비슷하였지만, 저온 8, 12주 처리는 2주차에 각각 68.7, 59.5%로 급격하게 발아율이 높아진 것을 볼 수 있었다. 하지만 4주차에는 모든 처리구에서 약 90% 이상 높게 발아하였으며, 통계적 유의성을 보이지 않았다. 위 결과를 통해 저온층적처리는 저온 처리기간이 길수록 발아소요기간을 단축하는 효과를 볼 수 있었다.
고 찰
종자는 내부 배의 형태에 따라 크게 기저형(basal), 지협형(peripheral) 및 중축형(axile)의 세 가지로 분류된다(Martin, 1946). 중축형은 배가 작은 것부터 큰 것까지 다양하게 보고하였다. 뽕나무과(Moraceae)의 종자 내부 형태는 배가 주걱모양으로 길게 구부려진 모양으로 보고된 바 있다(Barbour et al., 2008). 애기닥나무 종자의 배 형태도 중축형에서 구부러진 굽은(bent) 형태로 판단되어 기존 연구와 일치하였다(Fig. 2).
종피나 과피의 불투수성으로 인하여 야기된 휴면은 물리적 휴면(PY)으로 분류하는데(Baskin et al., 2000), 종자 초기 무게의 약 20% 이상 수분을 흡수하면 투수성을 갖는다고 판단할 수 있다(Baskin and Baskin, 2003). 애기닥나무 종자는 24시간 만에 75.1%의 수분흡수율을 나타내어 투수성을 갖기 때문에 물리적 휴면(PY)이 없다고 판단되었다(Fig. 3). 미숙배로 분류된 종자는 발아하기 전에 배가 일정 크기 이상 자라야 하는데, 적절한 조건에서 30일 이내에 배의 신장과 발아가 이루어질 경우 이를 형태적 휴면(MD)이라 한다(Baskin and Baskin, 1998). 하지만 애기닥나무 종자의 배는 완전히 발달된 배를 가지고 있기 때문에 형태적 휴면(MD)이 없다고 판단되었다. Baskin and Baskin (1998)은 종자가 탈리되는 시점에 배가 완전히 자라 있으나 발아가 30일 이상 지연될 경우 이를 생리적 휴면(PD)으로 분류하였다. 기존 문헌에서 Broussonetia papyrifera의 종자는 30일 이전에 35/15℃ 온도조건에서 발아를 하였다고 보고되어 휴면유형은 non dormancy를 가지고 있었다(Baskin and Baskin, 1998). 하지만 애기닥나무 종자는 30일 이전에 발아를 하였지만, 온도조건에 따라 발아가 지연되는 conditional dormancy 유형을 가지고 있다고 판단되었다(Fig. 4).
Baskin and Baskin (1998, 2004)은 배양 온도에 따라 발아가 지연되는 conditional-dormancy type에 대해서 보고한 바 있다. Conditional dormancy 휴면유형은 종자가 좁은 범위의 발아 적온에서만 발아하는 특성을 가지고 있다(Hartmann et al., 2002). Jones et al. (1997)은 Picea sitchensis 종자가 5℃에서 98%의 발아율을 보인 반면, 30℃에서 23%의 발아율을 보여 conditional dormancy 유형에 대해 보고하였다. Picea sitichensis 종은 10℃에서 휴면을 가졌지만, 4℃에서 12~14주 동안의 건조저장을 통해 휴면을 타파시켰다(Jones et al., 1988) Copete et al. (2009)은 이베리아 반도 2종의 conditional dormancy 유형을 보고하였는데, 상대적 저온(5℃, 15/4℃, 20/7℃)에서는 95%까지 발아를 하였지만, 상대적 고온(28/14℃, 32/18℃)에서는 발아하지 않는 특성을 가지고 있었다. 하지만 약 1~2개월의 건조저장을 통해 고온(28/14℃, 32/18℃)에서의 발아율을 약 90%이상 증가시켰다. 이처럼 conditional dormancy를 가진 유형은 저온과 고온의 특정 온도에서 휴면을 가지는 경우가 있으며, 좁은 범위의 발아 적온에서만 발아하는 특성을 가지고 있다고 볼 수 있다. 또한 Solanum mauritianum의 종자는 30/15℃에서 발아율은 80%였지만, 20℃에서의 발아율은 8%였다. 하지만 GA3를 통해 20℃에서의 발아율을 78%까지 높여 conditional domancy에 대해 보고하였다(Campbell et al., 1992). 애기닥나무 종자는 25/15℃ 온도조건에서 최종 발아율이 91.7%였지만, 15/6℃와 20/10℃의 경우 최종 발아율이 42.5, 44.4%로 절반 이상 발아율이 낮았다(Fig. 4). 하지만 GA3 처리를 통해 발아 소요기간을 줄이고, 상대적으로 낮은 온도조건에서 발아율을 높이는 것을 확인하여 이를 통해 conditional dormancy 유형으로 판단할 수 있었다(Fig. 5).
결론적으로, 애기닥나무 종자는 25/15℃의 온도조건에서 높은 발아율을 보였으나, 20℃ 이하의 상대적으로 낮은 온도조건에서 발아율이 급격하게 낮아지는 경향을 보였다. 위 결과를 통해 conditional dormancy로 판단할 수 있는데 GA3처리를 통하여 67.3%까지 상대적으로 낮은 온도조건에서 발아율을 높일 수 있었다. 이러한 결과를 통해 애기닥나무의 발아특성을 파악하였고, 대량증식 체계를 구축하여 기능성 소재 자원으로 쓰일 수 있는 기초자료를 확보하였다. 추후 저온층적처리를 통해서도 상대적으로 낮은 온도조건에서 발아율을 높일 수 있는지 추가적인 연구가 필요하다.
적 요
본 연구는 동아시아 특산식물 애기닥나무의 자원식물로서 활용을 위한 대량증식법 개발을 위해 실시하였다. 종자의 발아특성을 알아보기 위해 광·온도조건, GA3 처리, 저온층적처리를 실시하였다. 온도조건 실험에서 4, 15/6, 20/10, 25/15℃에서 발아율은 각각 0, 42.5, 44.4, 91.7%였다. 따라서 애기닥나무 종자의 발아 최적온도는 25/15℃였다. 온도조건에 따라 발아가 제한되었지만, GA3 처리를 통해 상대적으로 낮은 온도조건에서도 발아율을 높일 수 있었다. 저온층적처리를 통해 발아소요일수를 단축할 수 있었으며. 이를 통해 애기닥나무 종자를 conditional dormancy 유형으로 분류하였다. 본 연구결과를 통하여 적정 발아 온도조건은 25/15℃이며, 상대적으로 낮은 온도조건에서도 GA3처리로 종자기반 대량증식이 가능하며, 향후 산업적인 활용에 도움이 될 것이라 판단된다.
