

서 언
빌레나무(Maesa japonica (Thunb.) Moritzi & Zoll.)는 자금우과(Myrsinaceae)에 속하며 높이 1-1.5 m의 덩굴성 상록활엽수종으로 개화기는 4-5월이고 화서는 총상화서로 길이 1.5- 2.5 ㎝이다. 한 화서 당 10-27개의 꽃이 달리며, 꽃 색은 녹색으로 가장자리는 흰색이다(Moon et al., 2006). 빌레나무의 분포 지역은 중국, 대만, 일본 및 베트남 북부 등이며(Hyun et al., 2019), 국내에서는 2006년 제주도 곶자왈 지역에서 최초 발견되었다(Moon et al., 2006). 국내 분포 지역이 매우 협소하고 개체수가 희귀하여 제주지역 적색목록종으로 지정되었으며(Kim et al., 2008; 2011), 제주고사리삼(Mankyua chejuense Sun), 창일엽(Microsorium superficiale (Blume) Ching), 밤일엽(Neocheiropteris ensata (Blume) Ching), 제주방울란(Habenaria chejuensis Lee & Lee) 등과 더불어 곶자왈 한정 분포종으로 선정되었다(Hyun et al., 2019).
빌레나무는 제주도 서부 곶자왈 지역 내 종가시나무(Quercus glauca Thunb.), 팽나무(Celtis sinensis Pers.) 등이 수관을 우점하는 어두운 환경에서 자생한다(Song, 2011). 광 환경은 식물의 대사 활동에 많은 영향을 미치는 요인 중 하나로 수목의 광보상점, 광포화점, 최대 광합성속도, 호흡 작용, 엽록소 함량, 엽록소 비율 등에 매우 큰 영향을 미친다(Kim and Lee, 2001; Kwon et al., 2000; Lee, 2015). 광량이 부족한 환경에서 내음성이 약한 수종은 엽록소 함량 감소 및 광합성의 저하로 생장이 저조하지만 내음성이 상대적으로 강한 수종은 생리·형태적 반응을 보이며 광량이 부족한 환경에 적응하는 경향을 보인다(Cho et al., 2007, 2008; Kim and Lee, 2001). 반면 음지 및 반음지성 식물은 강한 빛에 의해서 잎의 엽록소가 파괴되고 황화현상이 나타나며 잎이 부패하거나 고사에 이르기도 한다(Kim et al., 2012; Oh et al., 2021). 따라서 수목의 건전한 생육 및 생존을 위해서는 수종별로 적정 수준의 광량이 요구된다.
한편, 최근 온난화의 영향으로 수목 북방한계선이 높아짐에 따라 난대활엽수종의 생육가능 지역이 확대되고 있다(Kim, 2010; Kwon et al., 2008). 이에 따라 경관 및 복원을 위한 조림, 약용, 실내조경 등 다양한 분야에서의 난대활엽수종의 수요가 증가하고 있다(Kim, 2010). 특히 빌레나무는 음지에서 생육하는 특성을 고려하면, 실외 조림에만 국한되지 않고 실내 조경수로도 활용될 가능성이 크다. 또한 벤조퀴논 유도체(Ogawa and Natori, 1965; 1968), 사포닌 성분(Foubert et al., 2009; Ngo et al., 2019), 항산화 물질, 항미생물 활성(Kim, 2014) 등 여러 유용한 성분에 관한 보고가 있어 유용 수종으로의 잠재력도 크다.
빌레나무에 관한 선행연구는 앞서 언급한 것처럼 유용성분들에 관한 연구가 주로 보고되고 있고 개체발생학적, 분류학적, 형태학적 연구(Caris et al., 2000; Ma and Saunders, 2003; Utteridge and Saunders, 2006)가 보고되고 있다. 그러나 빌레나무의 생육 및 증식에 관한 선행연구는 Kwon et al. (2020)이 보고한 온도, 지베렐린 산, 질산칼륨 처리에 따른 종자 발아율 연구를 제외하고는 보고된 바가 없어 관련 연구가 미진한 실정이다. 따라서 본 연구는 차광 처리에 따른 빌레나무의 개화 및 생장 반응을 조사하여 희귀 수종인 빌레나무 생육에 알맞은 최적의 광조건을 규명하고, 이를 통하여 빌레나무의 건전한 생육 및 유무성증식을 통한 보급 증대에 기여하고자 한다.
재료 및 방법
연구 재료 및 실험구 처리
본 연구는 서귀포시 상효동에 위치한 난대아열대산림연구소 내 양묘장에서 수행되었다. 공시수종인 빌레나무는 2019년 7월 연구시험림에서 채집하여 연구소 내 온실에서 삽목으로 증식시킨 1년생 개체를 사용하였다. 시판되는 차광막(차광률 35%, 55%, 75%, PYUNG HWA INDUSTRY, Korea)을 이용하여 세 조건의 차광 처리구를 설정하였고, 비교구로 실외에 비차광 처리구를 설정하였다. 2020년 5월 각 처리구에 약 130-160본의 빌레나무를 25 ㎝ 간격으로 이식하였다. 가장자리에 있는 묘목을 제외하고 27-30본의 묘목을 한 조사구로 설정하였고, 각 처리구마다 조사구를 3반복하였다. 한편, 2021년 1-2월 비차광 처리구의 모든 묘목의 지상부가 고사하였다. 2021년 2-3월 대기온도를 측정하기 위하여 차광 처리구와 빌레나무 자생지에 데이터 로거(EL-USB-2-LCD+, LASCAR Elctronics, Hong Kong)를 설치하였다. 차광 처리구의 토성은 사질양토이고, 토양 특성의 평균값은 pH 5.6, 유기물 10.7%, 전질소 0.38%, 양이온치환능력 25.4 cmolc/kg, 전기전도도 0.15 dS/m 등이며, 처리구간 유의한 차이는 나타나지 않았다(p > 0.05).
개화 관찰
각 조사구의 모든 묘목의 개화를 6단계(Fig. 1; 1단계: 화서 신장, 2단계: 개화 시작, 3단계: 약 10-40% 꽃 개화, 4단계: 40-70% 꽃 개화, 5단계: 70% 이상 꽃 개화, 6단계: 꽃 전부 짐)로 나누어 2021년 1-5월에 묘목의 개화 단계를 측정하였다. 개화 전에는 주 1회, 개화 시작 후에는 주 2회, 총 26회 관찰하였고, 각 처리에 따른 개화 시기와 개화율, 화서의 수를 측정하였다.

생장 측정
각 조사구의 모든 묘목을 대상으로 2020년 5월에 묘고와 잎의 수를 측정하였고, 처리에 따른 통계적인 차이는 나타나지 않았다(p > 0.05). 2021년 3월에 묘고, 잎의 수, 가지의 수를 측정하였다. 묘고는 가장 긴 가지의 길이를 측정하였다. 묘목의 생물량을 측정하기 위해 2021년 3월에 처리구별로 묘목을 3본씩 굴취하여 잎, 줄기, 뿌리로 분리하였다. 잎은 이미지 분석 프로그램(ImageJ, National Institutes of Health, USA)를 이용하여 면적을 측정하였고, 뿌리, 줄기, 잎을 65℃에서 건조한 후 건중량을 측정하였다. 이를 이용하여 지상부 대비 뿌리 건중량 비율(R/S; root weight to shoot weight ratio), 총 건중량 대비 잎의 건중량 비율(LWR; leaf weight ratio), 잎의 건중량 당 면적 비율(SLA; specific leaf area)을 계산하였다.
통계분석
차광 처리에 의한 묘고, 잎의 수, 가지의 수, 건중량, 엽면적 및 각 비율 차이의 유의성은 일원분산분석을 이용하였고, Duncan의 다중비교법으로 사후검정을 하였다. 모든 통계분석은 SPSS version 28 (IBM, USA)을 이용하였다.
결과 및 고찰
차광처리에 따른 개화 반응
빌레나무는 차광 처리 별로(35%, 55%, 75%) 각각 평균 3월 24일, 3월 24일, 3월 29일에 개화가 시작되었으며, 차광률 증가에 따라 개화가 지연되는 경향이 나타났다(Fig. 2). 개화 기간(일)은 차광 처리 별로(35%, 55%, 75%) 평균 23.2 ± 2.0, 26.6 ± 0.9, 24.9 ± 2.3이며, 처리에 따른 유의한 차이를 보이지 않았다(p > 0.05). 한편, 차광 처리 별로(35%, 55%, 75%) 한 개체 당 화서의 수(개)는 3.9 ± 1.3, 4.7 ± 0.7, 2.7 ± 0.2이고(Fig. 3a), 처리구 당 개화한 개체의 비율(%)은 27.3 ± 3.6, 31.6 ± 9.2, 21.5 ± 3.9 등으로(Fig. 3b) 55% 처리구에서 가장 많은 개체가 개화하고 개체당 화서수도 높은 경향을 보였다.
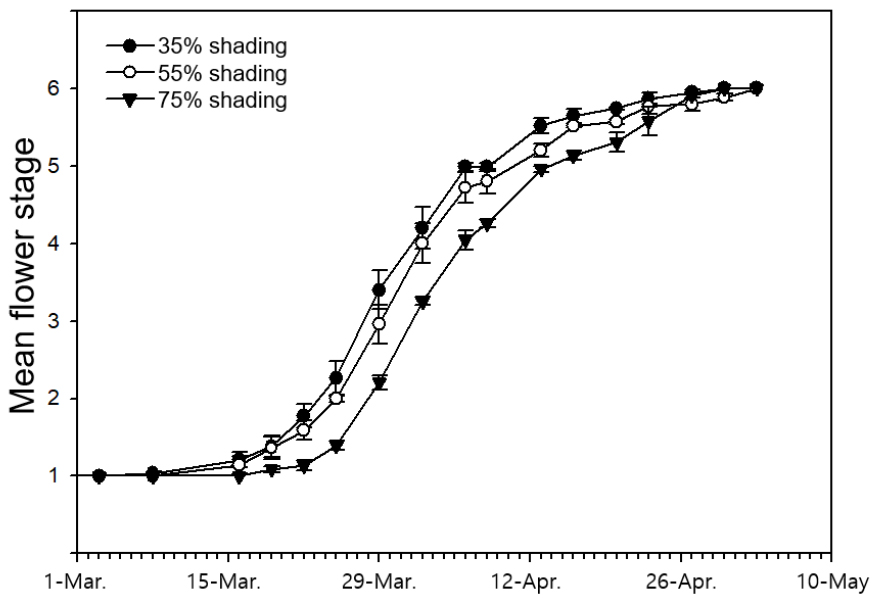
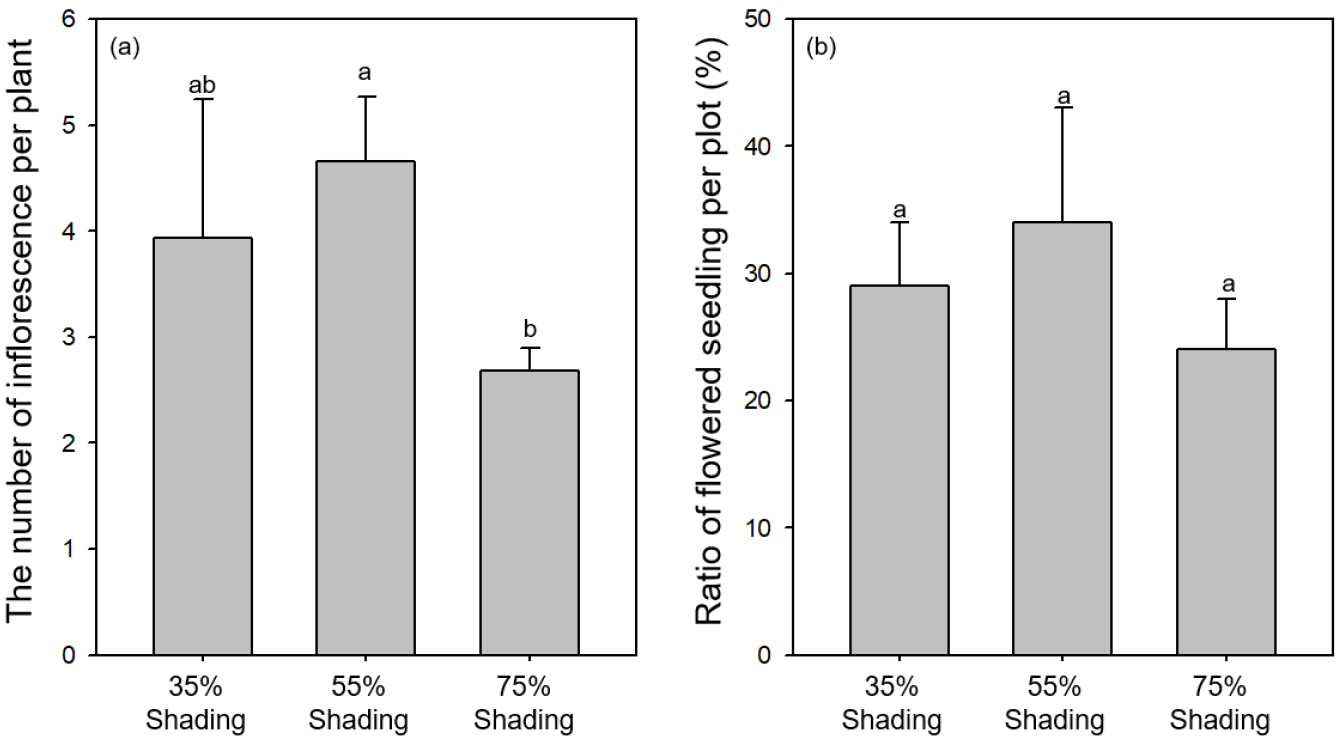
Fig. 3.
The number of inflorescence per plant (a) and ratio of flowered seedling per plot (b) of M. japonica under artificial shading treatments. Error bars indicate the standard errors of the mean. Values with the different letter (a-b) indicate significant difference among three treatments (p < 0.05).
개화 시기는 차광 처리에 따라 촉진되거나(Seo et al., 2002) 변화가 없는(Chung and Yoo, 1997) 등 수종, 처리 기간, 환경 등에 따라 다양한 연구 결과가 보고되고 있다(Cheong et al., 2005; Nam et al., 2012). 그 중 유색칼라(Zantedeschia elliottiana (W.Watson) Engl.)는 차광 처리에 따라 10-59일 개화가 지연되고(Nam et al., 2012), 새우난초(Calanthe discolor Lindl.)는 차광 처리별로 1-9일이 지연되어(Kim et al., 2012) 차광률 증가에 따라 개화 시기가 지연되는 것으로 나타났으며, 본 연구 결과도 이와 같은 경향을 보였다. 한편, 차광 처리에 따른 개화 기간은 중간 정도의 차광 처리에서 가장 긴 것으로 보고되며(Son and Chae, 2003, Seo et al., 2002), 본 연구 결과도 통계적 유의성은 없으나 이와 동일한 경향성을 보였다.
차광 처리에 따른 개화율과 화서의 수도 개화 시기와 마찬가지로 다양한 결과가 보고되고 있다. 차광 처리에 따라 소화(floret)수가 감소하거나 개화 전 꽃이 고사하는 경우가 보고되었으며(Hong et al., 2006; Jung and Choi, 1999), 이와 반대로 차광률이 증가할수록 개체 당 화서의 수가 증가하는 사례도 보고되었다(Chung and Yoo, 1997). 그러나 앞선 연구와 다르게 중간 정도의 차광 처리가 개화에 유리한 경향도 보고되고 있다. 타래난초(Spiranthes sinensis (Pers.) Ames)는 30%, 50%, 70% 차광 처리구 중 50% 차광 처리에서 가장 높은 소화수를 보였고(Son and Chae, 2003), 유색칼라 또한 50% 차광에서 절화수가 가장 많았다(Nam et al., 2012). 새우난초는 포기당 꽃대의 수가 비차광 처리에서 가장 적고 58% 차광 처리에서 가장 높았으며, 화서 당 소화수는 71%에서 가장 높은 것으로 나타났다(Kim et al., 2012). 화아를 유도하는 요인은 식물의 영양상태, 식물 호르몬, 광 조건, 온도 조건 등이 있는 것으로 알려져있다(Kang et al., 2009). 식물은 광합성 산물을 영양생장에 우선적으로 분배하여 일정한 수준의 잎을 먼저 생성하고 그 후에 꽃을 생성한다(Obeso, 2002; Wang et al., 2020). 즉, 개화 촉진을 위하여 양호한 영양상태가 필요하며, 과도한 광량의 감소는 광합성 산물의 부족을 야기하여 꽃의 발육을 지연시키거나 정지시키게 된다(Bodson, 1983). 이러한 이유로 본 연구에서 차광률 증가에 따라 개화가 지연되고, 75% 차광 처리구의 소화수가 유의하게 감소한 것으로 판단된다.
차광 처리에 따른 생장 반응
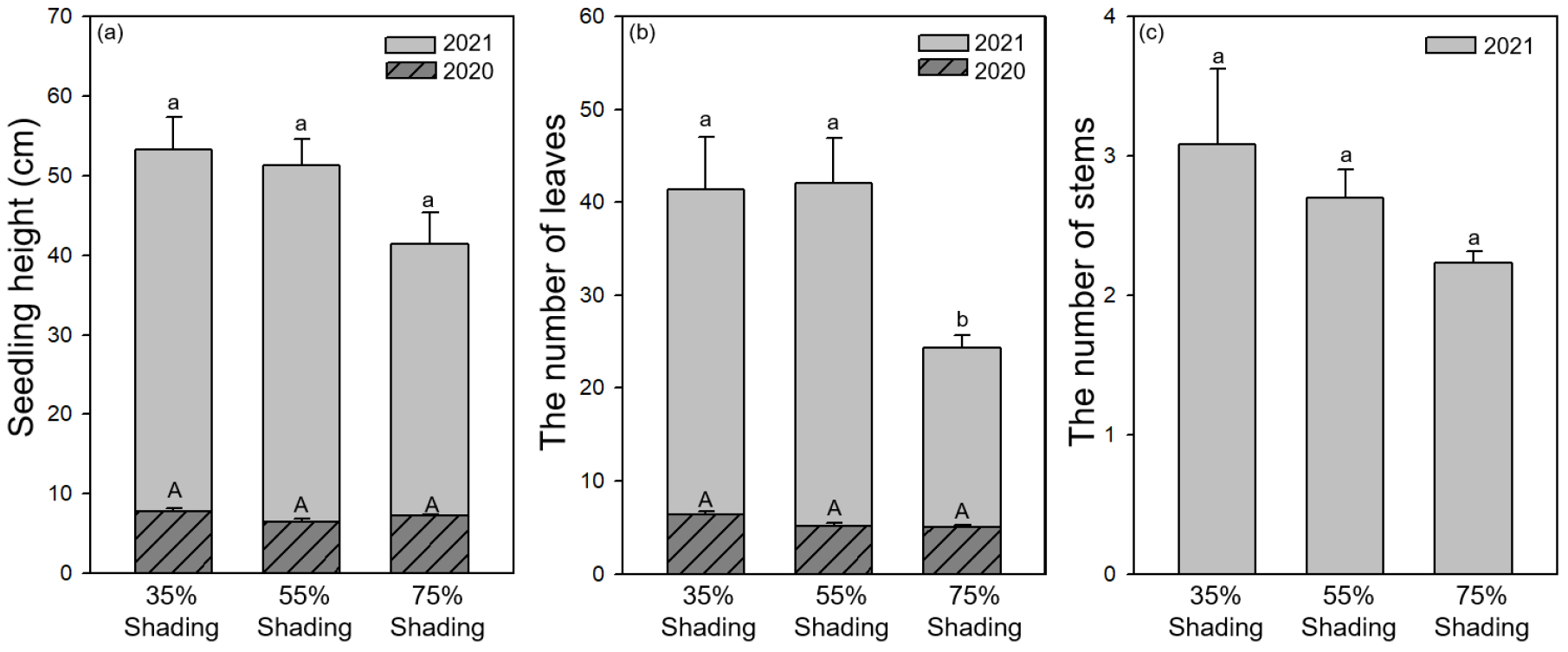
빌레나무의 평균 묘고(㎝)는 차광 처리 별로(35%, 55%, 75%) 각각 53.2 ± 4.2, 51.3 ± 3.3, 41.4 ± 4.0 순으로 나타났고, 차광률이 증가함에 따라 묘고가 감소하는 경향을 보였으나 처리 간의 유의성은 나타나지 않았다(Fig. 4a). 잎의 수(개)는 차광 처리 별로(35%, 55%, 75%) 각각 41.3 ± 5.6, 42.4 ± 4.8, 24.4 ± 1.3 등이며 75% 차광 처리구의 잎의 수는 다른 처리구 대비 유의하게 적은 것(p < 0.05)으로 나타났다(Fig. 4b). 차광 처리 별로(35%, 55%, 75%) 줄기의 수(개)는 각각 3.1 ± 0.5, 2.7 ± 0.2, 2.2 ± 0.1 순으로 차광률 증가에 따라 감소하는 경향을 보였으나 통계적으로 유의한 차이를 보이지 않았다(Fig. 4c).
빌레나무 묘목의 생물량을 분석한 결과, 전체, 지상부, 잎, 줄기 및 지하부 생물량 모두 35% 차광 처리구에서 가장 높으며, 차광률이 증가함에 따라 전체 및 부위별 건중량이 감소하는 경향을 보였다(Table 1). 35% 차광 처리구의 생물량은 75% 차광 처리구의 생물량보다 전체는 2.0배, 지상부, 잎, 줄기는 2.1배, 지하부는 1.7배 높은 것으로 나타났으며, 통계적인 유의성을 보였다(p < 0.05). 총 엽면적은 35% 차광 처리구에서 75% 차광 처리구보다 2.0배 높았으며(p < 0.1) SLA와 LWR은 처리에 따른 유의성이 나타나지 않았다(Table 2). 한편, 비차광 처리구의 모든 묘목은 2021년 1월 지상부가 전부 고사하였다.
Table 1.
Biomass and root to shoot ratio (R/S) of M. japonica seedlings with artificial shading in March 2021
| 35% shading | 55% shading | 75% shading | |
| Total (g) | 21.29 " 2.36az | 18.35 " 2.39a | 10.66 " 1.19b |
| Aboveground (g) | 16.84 " 2.04a | 14.76 " 1.85a | 8.10 " 1.30b |
| Leaf (g) | 11.29 " 1.52a | 9.48 " 1.20ab | 5.48 " 0.88b |
| Stem (g) | 5.55 " 0.52a | 5.27 " 0.66a | 2.61 " 0.43b |
| Root (g) | 4.45 " 0.40a | 3.60 " 0.55ab | 2.56 " 0.21b |
| R/S ratio | 0.27 " 0.02a | 0.26 " 0.00a | 0.35 " 0.07a |
Table 2.
Total leaf area, specific leaf area (SLA) and leaf weight ratio (LWR) of M. japonica seedlings with artificial shading in March 2021
| 35% shading | 55% shading | 75% shading | |
| Total leaf area (㎠) | 1,123.41 " 214.98az | 880.08 " 101.75ab | 575.24 " 78.82b |
| SLA (㎠·g-1) | 98.39 " 7.42a | 93.10 " 1.79a | 104.59 " 5.08a |
| LWR (g·g-1) | 0.52 " 0.01a | 0.51 " 0.00a | 0.50 " 0.02a |
차광 처리에 따른 식물의 생장 반응은 수종과 처리기간에 따라 다양하게 보고되고 있다. 고로쇠나무(Acer mono Maxim), 물푸레나무(Fraxinus rhynchophylla Hance), 층층나무(Cornus controversa Hemsl.), 박달나무(Betula schmidtii Regel) 등 5개 활엽수종을 대상으로 한 4단계 수준의 차광 처리 연구 결과에 따르면 처리 기간이 길어질수록 차광 처리의 영향이 강하게 나타난다고 하였으며, 고로쇠나무를 제외한 4개 수종은 차광률이 감소할수록 상대생장률과 생물량이 증가한다고 보고하였다(Cho et al., 2008). 그러나 묘목 단계에서 내음성이 강한 식물은 광도가 높은 환경에서 광 스트레스로 인해 생장이 저조하고, 동시에 적정 수준 이하의 광도에서는 생장률이 크게 감소하여 적절한 수준의 차광 처리가 필요한 것으로 알려져 있다(Choi et al., 2002; Kobe et al., 1995). 고로쇠나무, 죽절초(Sarcandra glabra (Thunb.) Nakai), 백량금(Ardisia crenata Sims), 눈측백(Thuja koraiensis Nakai), 굴거리나무(Daphniphyllum macropodum Miq.) 등 내음성이 있는 것으로 알려진 수종들은 중간 정도의 차광 처리에서 가장 높은 생장과 생물량을 보이는 것으로 보고되었으며(Bae et al., 2016; Cho et al. 2008; Hong et al., 2000; Song et al., 2015), 본 연구 결과도 이와 유사하였다.
광량이 부족한 환경에서는 광합성 산물의 절대적인 양이 감소하지만(Cho et al., 2001; 2008) 광 경쟁에서 유리한 위치를 차지하고자 수고 생장으로 분배되는 광합성 산물이 상대적으로 증가한다(Kozlowski et al., 1991). 본 연구 결과 빌레나무의 지상부 생물량과 잎의 수는 75% 차광 처리구에서 35% 차광 처리구 대비 각각 2배, 1.7배 낮아 처리에 따른 뚜렷한 차이를 보였는데 묘고는 유의한 차이를 보이지 않았다. 이는 75% 차광 처리구의 빌레나무가 상대적으로 광량이 적은 환경에 적응하기 위하여 수고 생장에 양분을 더 분배한 결과인 것으로 판단된다.
한편, 생식생장과 영양생장은 식물체 내의 탄소와 질소 비율, 양분 경쟁 등으로 한 종류의 생장이 더욱 왕성하게 나타나거나 서로 부의 상관관계를 나타내기도 한다(Kang et al., 2009; Koenig and Knops, 1998). 새우난초의 경우 생물량은 58% 차광 처리에서, 꽃의 수는 71%, 80% 차광 처리에서 높게 나타났고(Kim et al., 2012), 본 연구에서도 빌레나무의 영양생장과 생식생장이 각기 다른 차광 수준에서 왕성한 것으로 나타났다.
총 엽면적은 35% 차광 처리구와 75% 차광 처리구 간 1.9배 차이로 통계적 유의성을 보였으며 잎의 수와 상관관계가 있는 것으로 나타났다(r = 0.88). 일반적으로 광량이 감소하면 암환경에 적응하기 위하여 SLA나 LWR이 증가하는 경향이 나타나지만 수종에 따라 감소하거나 일정한 경향이 없는 등 다양하게 보고되고 있다(Cho et al., 2008, Poorter, 1999). 본 연구 결과 차광 처리에 따른 SLA와 LWR의 차이는 나타나지 않았으나 빌레나무가 다른 활엽수종에 비하여 SLA가 낮고 LWR이 높은 것을 확인할 수 있었다. 이는 빌레나무가 덩굴성 식물이기 때문인 것으로 판단되며, 덩굴 식물은 잎 생산에 비 덩굴성 수종보다 많은 양분을 분배하여 기저면적 대비 잎의 비율이 높게 나타난다(Paul and Yavitt, 2011).
한편, 비차광 처리구의 모든 묘목에서 엽소(leaf burn)현상이 관측되었고, 2021년 1월 지상부가 고사하였다. 이는 광 스트레스의 영향을 받은 것으로 판단된다. 묘목 단계에서 내음성이 강한 수종은 강한 광 환경에서 생장이 저조한 것으로 보고되고 있다(Kobe et al., 1995). 광이 강한 환경에서는 엽록소 분해 작용으로 잎의 색이 퇴색될 수 있고, 세포액의 산도 감소가 철분의 이동을 방해하여 황화현상이 나타나며, 체내의 수분 평형을 유지하기 위하여 기공을 닫아 광합성을 중지하기도 하는 것으로 알려져 있다(Kim et al., 1997). Seo et al. (2002)은 은방울꽃(Convallaria keiskei Miq.)의 엽소율이 비차광 처리에서 73.8%로 가장 높았으며, 차광률이 증가할수록 감소한다고 보고하였다. 또한 빌레나무와 유사한 환경에서 자생하는 새우난초에 대한 차광 연구 결과, 광량이 증가하면 엽록소 함량이 매우 낮아지고, 황화현상이 심하게 나타나며, 상단부의 많은 부분이 고사하는 증상이 있는 것으로 나타났다(Kim et al., 2012). 비차광 처리구의 빌레나무도 광 스트레스를 받았을 것으로 보이며, 이는 생장에 부정적인 영향을 주었을 것으로 판단된다. 한편, 수목에 스트레스를 주어 고사에 이르게 하는 요인은 광, 온도, 수분, 바람, 양분 부족 등으로 다양하며(Franklin et al., 1987; Lee, 2015; Woodward et al., 2004), 특히 난대활엽수종은 최한월 기온의 영향을 많이 받는 것으로 알려져 있다(Koo et al., 2001). 또한 본 연구에서 빌레나무의 생장 반응이 35%와 75% 차광 처리 간 유의한 차이를 보였지만 35%와 55% 차광 처리 간에는 명확한 차이가 나타나지 않았다. 따라서 앞으로 온도, 수분 등의 여러 환경적 요인과 0-55% 사이의 차광 조건을 추가하여 연구를 지속할 필요가 있을 것으로 판단된다.
적 요
본 연구는 제주도 곶자왈 지역에 자생하는 빌레나무를 대상으로 차광 처리를 통해 세 가지 수준의 차광 조건(35%, 55%, 75%)에 따른 개화 및 생장 특성을 분석하였다. 차광률 증가에 따라 개화 시기가 지연되었고, 55% 차광 처리구에서 가장 많은 개체가 개화하였으며, 개체당 화서의 수도 높게 나타났다. 차광률이 증가할수록 묘고가 감소하였고 잎과 줄기의 수는 75% 차광 처리에서 가장 낮은 것으로 나타났다. 전체 및 부위별 생물량도 차광률 증가에 따라 감소하였으며, 35% 차광 처리구에서 75% 차광 처리구 대비 지상부 생물량 2.1배, 지하부 생물량 1.7배, 총 엽면적 2.0배 높은 것으로 나타났다. 종합적으로 고려하면 빌레나무는 건전한 생육을 위하여 차광 처리가 필요하며 개화는 55% 차광 수준, 생장은 35% 차광 수준에서 가장 양호한 것으로 판단된다.
