

Introduction
Materials and Methods
Plant material and in vitro propagation
Cold acclimatization of donor plantlets
Shoot tip isolation
Loading and dehydration procedure for cryoprotection
Ultrarapid cooling by liquid nitrogen
Thawing and recovery procedure
Assessment of plant viability and recovery
Statistical analysis
Results and Discussion
Recovery rate in routine cryopreservation
Protocol optimization based on initial cryopreservation results
The utilization of larger apical shoot tip dimensions
Duration of dehydration
Variation in cold acclimation procedure
Carbon source adjustment for donor plant optimization
Introduction
As one of the top three staple crops, potatoes play a crucial role in global food security and poverty alleviation (Devaux et al., 2014; Niino and Arizaga, 2015). With a higher harvest index compared to that of grains, potatoes are cultivated on 20.7 million hectares worldwide, yielding 437 million tons annually, with an average yield of 20.9 tons per hectare. Beyond their role as a food crop, more than 4,500 varieties of Solanum tuberosum ssp. tuberosum are grown globally, serving as essential resources for industries such as animal feed, pharmaceuticals, textiles, and paper production (Nagel et al., 2022).
Today, ‘Solanum tuberosum’ refers to improved potato varieties that have undergone intensive breeding over the past 200 years. Unlike wild potatoes, which can be preserved using botanical seeds, these cultivated varieties must be propagated and maintained clonally through vegetative tissues to preserve their uniform characteristics (George et al., 2017; Niino and Arizaga, 2015). Therefore, various conservation strategies have been employed, including in situ preservation, ex situ collection preservation in fields or greenhouses, preservation of vegetative propagules (e.g., tuber collections), in vitro preservation, and cryopreservation (Vollmer et al., 2022). Worldwide, 82,293 potato accessions are conserved across 89 institutions and 4 international or regional centers in 59 countries. However, more than 50% of the world’s potato genetic resources are concentrated in key locations such as Germany, France, India, Russia, the United States, and the International Potato Center (CIP) in Peru (Nagel et al., 2022). Despite these efforts, many genebanks in Latin America and parts of Europe have traditionally preserved their entire potato collections in fields, leaving them vulnerable to environmental risks such as pathogen and pest infestations, which increase the likelihood of genetic drift and resource loss (Nagel et al., 2022; Vollmer et al., 2022). Initially, CIP also relied on ex situ preservation via annual tuber regeneration; however, the introduction of in vitro technologies in the 1980s enabled the conservation of microtubers or plantlets under low-temperature conditions. Although in vitro preservation offers the advantage of sterile storage, it requires regular subculturing and carries the risk of somaclonal variation over time. Moreover, repeated regeneration process increases the potential for human error or biological changes, which can accumulate and compromise the integrity of the preserved materials (Anglin et al., 2021).
Cryopreservation offers an effective alternative for mitigating numerous limitations. This method, which involves storing biological samples in liquid nitrogen at -196°C, securely preserves clonal crops' germplasm from multiple risks while ensuring cost and space efficiency (Vollmer et al., 2021). Numerous studies have demonstrated that cryopreserved plant materials retain viability over extended periods and can regenerate into healthy plants after thawing (Song et al., 2023). Moreover, genetic variations that may arise during long-term subculturing can be effectively minimized with the application of optimized cryopreservation protocols (Niino and Arizaga, 2015). Furthermore, research conducted by the Leibniz Institute of Plant Genetics and Crop Plant Research (IPK) in Germany has shown that the cost of cryopreserving potatoes is $6.4–$12 per sample, significantly lower than the $49–$59 required for field collection. Despite higher initial costs, cryopreservation remains a cost-effective option for long-term conservation overtime (Keller, 2006; Keller et al., 2008; Schäfer-Menuhr et al., 1996). Since the late 1970s, various protocols for cryopreserving potato germplasm using in vitro shoot tips have been developed, including vitrification, encapsulation-vitrification, droplet-vitrification, and the V cryo-plate method (Halmagyi et al., 2004; Yi et al., 2012). These methods have undergone continuous refinement over the years (Benson et al., 2011; Panta et al., 2014; Wang et al., 2008). The International Potato Center (CIP), which houses the world’s largest cryopreserved potato collection, provides user-friendly droplet-vitrification protocols for large-scale cryobanking, covering more than 70 genotypes, including all cultivated varieties (Vollmer et al., 2021). These protocols, available through scientific publications and instructional videos on the CIP website, highlight the importance of cold acclimation prior to shoot tip excision, a critical step successfully applied to over 90% of tested accessions (Acker et al., 2017).
The Rural Development Administration (RDA) of Korea operates one of the potato cryobanks that preserve in vitro-grown shoot tips. According to database records, the National Agrobiodiversity Center (NAC) last cryopreserved 61 potato accessions in 2013. In 2022, the NAC resumed potato cryobanking efforts, prioritizing domestically developed cultivars. To execute this project, the NAC adopted the latest practical potato cryo-protocol developed by the International Potato Center (CIP) and plans to systematically implement the cryobanking of RDA-bred potato cultivars by 2026.
Implementing large-scale cryopreservation of plant genetic resources requires customizing standard protocols to accommodate diverse genotypes as a single protocol cannot ensure consistent survival and regeneration across accessions, even within the same crop This is particularly critical for genetically diverse crops like potatoes (Niino and Arizaga, 2015). For instance, the CIP potato cryobank reported that fewer than 10% of its entire collection responded poorly to the standard protocol. While optimizing protocols for these accessions is essential, it demands additional labor and funding, which can delay progress (Acker et al., 2017). Variability in survival rates is influenced by factors such as donor plant vigor, sensitivity to cryoprotectants (CPA), and the physiological condition of plant material (Engelmann, 2000).
This study applied CIP’s PVS2 droplet-vitrification protocol to eight RDA-bred potato varieties to establish cryo-collections for Korean cultivars. For cryopreservation-recalcitrant varieties (Zhang et al., 2023), the protocol was adjusted to improve regeneration rates. Optimization targeted varieties with statistically low post-thaw recovery rates, focusing on four factors: (1) carbon source concentration in the medium to enhance donor plant quality, (2) cold hardening duration, (3) shoot tip size, and (4) dehydration time.
Materials and Methods
To implement the cryobanking of Korean potato cultivars, the potato shoot tip cryopreservation protocol released by the International Potato Center was adapted to fit the conditions of the National Agrobiodiversity Center (NAC) in Korea. Regarding cultivars with low recovery rates under the protocol, the process was customized by: (1) adjusting sucrose concentration during in vitro multiplication (20.0-30.0g), (2) modifying tissue size (2.0-2.5 ㎜), and (3) adjusting the durations of cold hardening and (4) dehydration treatments. This operational protocol is available on the CIP website (https://cipotato.org/genebankcip/process/cryopreservation/).
Plant material and in vitro propagation
This study focuses on eight Korean potato cultivars developed by the Highland Agriculture Research Institute (HARI) of the National Institute of Crop Science (NICS), Rural Development Administration (RDA). These cultivars, classified as Solanum tuberosum subsp. tuberosum, are improved varieties with diverse traits, officially registered between 2009 and 2023 (Table 1). Cryopreservation materials included in vitro plantlets, with approximately 80 per accession provided by HARI to the NAC. Plantlets were screened for endophytic contamination, and only pathogen-free, healthy specimens were selected for mass propagation. For micropropagation, five apical segments (with 1-2 leaves) were planted in air-permeable culture vessels (Gaooze 0811C, Korea Scientific Technique Industry) and incubated at 24 ± 1°C under a 16-hour photoperiod with a light intensity of 100–140 μmol/ ㎡/s. Subculturing intervals ranged from 2 to 6 weeks depending on the growth rate of each genotype (Table 1). The propagation medium was comprised of solid Murashige and Skoog (MS) medium (Duchefa Biochemie, Netherlands) with vitamins, 20–30 g/L sucrose (genotype-dependent, ‘Jayoung’ cultivar), and 2.6 g/L phytagel. The ㏗ was adjusted to 5.8 before autoclaving at 121°C for 20 minutes.
Table 1.
List of eight cryopreserved potato cultivars developed by the RDA, Koreaz.
| IT No.y | Cultivarx | Scientific namew |
Subculture intervalv |
Biological statusw | Descriptionx |
| 245944 | Jayoung | Solanum tuberosum subsp. tuberosum | 3-4weeks |
Improved varieties |
Purple potato variety with high concentration anthocyanin content |
| 341132 | Dami | Solanum tuberosum subsp. tuberosum | 3-4weeks |
Improved varieties |
Potato variety with few tuber physical defects, high dry matter content |
| 341134 | Mangang | Solanum tuberosum subsp. tuberosum | 3-4weeks |
Improved varieties |
Potato Variety with good chipping quality and late blight resistance |
| - | Goldenball | Solanum tuberosum subsp. tuberosum | 3-4weeks |
Improved varieties | Potato Variety resistant to browning |
| 341135 | Arirang-1ho | Solanum tuberosum subsp. tuberosum | 2weeks |
Improved varieties |
Potato Variety with the High Temperature and Drought Resistances |
| 341136 | Arirang 2ho | Solanum tuberosum subsp. tuberosum | 5-6weeks |
Improved varieties | Potato Variety Tolerant to Drought and Heat Stress |
| 245942 | Hongyoung | Solanum tuberosum subsp. tuberosum | 2weeks |
Improved varieties |
Potato Variety with Red Skin and Flesh Color, and High Concentrations of Anthocyanins |
| 231908 | Haryeong | Solanum tuberosum subsp. tuberosum | 3-4weeks |
Improved varieties | Potato Variety resistant to late ‘blight and viruses |
zAll 8 cultivars were bred by the Highland Agriculture Research Institute (HARI), National Institute of Crop Science (NICS), RDA.
xThe English names and characteristics of the cultivars were cited from the website of the Korea Seed & Variety Service (KSVS).
wThe information was referenced from GMS website (Germplams Management System, https://genebank.rda.go.kr/).
Cold acclimatization of donor plantlets
During the final stage of propagation, uninodal stem segments, including axillary buds, were densely planted in culture vessel. The explants were incubated under normal temperature (24°C ± 1°C) and light conditions (16-hour photoperiod, 100-140 μmol/㎡/s) for 5–10 days, with apical shoot regeneration occurring from the axillary buds. Depending on the growth pattern of each cultivar, the plantlets underwent a primary cold hardening phase at 7 ± 1°C with a light intensity of 50 μmol/㎡/s for 10–20 days, followed by a secondary acclimatization phase at 4.5 ± 1°C for 1–4 days (CIP website https://cipotato.org/genebankcip/process/cryopreservation/, Fig. 1). For the ‘Hongyoung’ cultivar, the protocol was adjusted to include only the primary cold acclimation phase at 7°C.
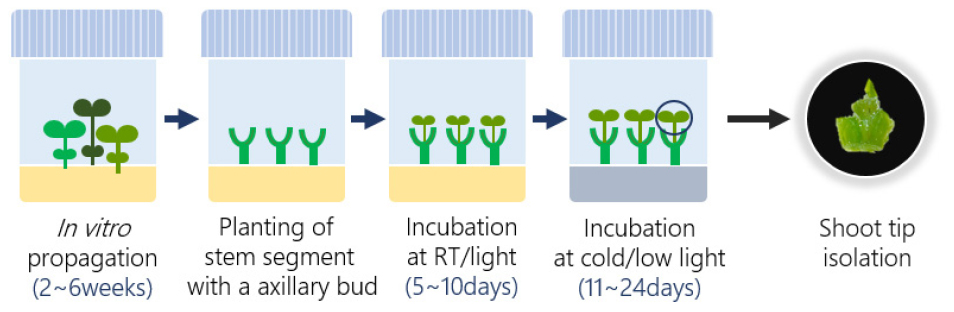
Shoot tip isolation
After a 2–3 week physiological acclimation in a cold chamber, shoot tips (1.5–2.5 ㎜) were excised from the apical region of in vitro-grown plantlets. The standard protocol describes shoot tips as measuring 0.8–1.2 ㎜ in length and 0.3–0.7 ㎜ in width, containing 2–4 leaf primordia, while also reporting that the length and width of shoot tips may vary depending on the potato genotype (Vollmer et al., 2022). Accordingly, in this study, the size of the explants was adjusted based on the post-thaw survival rate for each potato cultivar (Fig. 2, ‘Haryeong’ cultivar). For each cryopreservation trial, approximately 90-100 uniform shoot tips were prepared, with 10-20 shoot tips placed on sterile filter paper pieces, which served as carriers during the cryoprotection process (Vollmer et al., 2022; Fig. 3a).

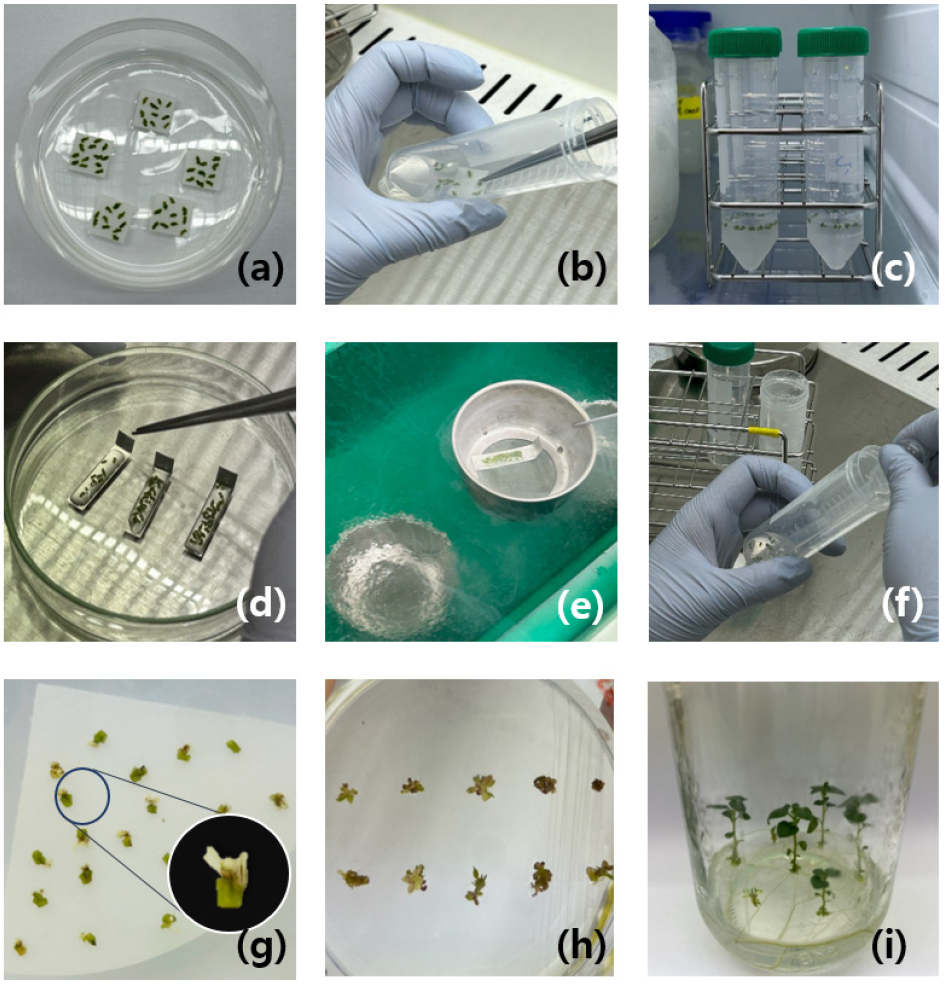
Fig. 3.
Cryopreservation procedure of korean potato varieties using the CIP protocol (Vollmer et al., 2021). (a) Isolated shoot tips placed on sterilized filter papers (size varies by cultivars, 1.5–2.0 ㎜). (b) Shoot tips immersed in a Falcon tube containing Loading Solution (LS). (c) Dehydration in a freezer below -10°C for 50 minutes. (d) Droplet formation of PVS2 containing shoot tips on an aluminum foil strip. (e) Ultra-rapid freezing of aluminum foil strips in liquid nitrogen for a minimum of 1 hour. (f) Thawing of shoot tips in Rewarming Solution for 20 minutes at RT. (g) Regeneration of surviving shoot tips incubated on recovery medium in darkness for 2 weeks. (h) Shoot tips at week 6 after subculturing on fresh MS medium under normal light conditions with observed leaves and stems (“Mangang”). (i) Plantlet at 7–8 weeks post-thaw culture with completed root formation (“Arirang-1ho”).
Loading and dehydration procedure for cryoprotection
This study circumvented the preculture procedure by implementing a cold acclimation strategy to induce the physiological adaptations necessary for enhanced freezing tolerance. Shoot tips excised from cold-acclimated plantlets were treated in a conical tube (Falcon tube, 50 mL, SPL, Korea) containing loading solution at room temperature for 20 minutes (Fig. 3b). The Loading Solution (LS) consisted of liquid MS medium (4.43 g/L) with vitamins, supplemented with 136.8 g/L sucrose and 147.4 mL/L glycerol. The ㏗ of the solution was adjusted to 5.8 prior to filter sterilization (pore size: 1.0 ㎛). Subsequently, the LS was removed from the tube and replaced with precooled Plant Vitrification Solution 2 (PVS2). The shoot tips were then subjected to dehydration in a freezer set at -18°C for 30 to 70 minutes (Fig. 3c, ‘Hongyoung’ cultivar). The dehydration duration in the standard protocol is 50 minutes; however, the treatment time was adjusted as needed based on genotype, sample size, and sensitivity to the cryoprotectant.
PVS2 is formulated with liquid MS medium supplemented with 240.0 mL/L glycerol, 134.8 mL/L ethylene glycol (324558, Merck Sigma-Aldrich®), 136.4 mL/L dimethyl sulfoxide (D4540, Merck Sigma-Aldrich®), and 136.8 g/L sucrose. The ㏗ was precisely adjusted to 5.8. This dehydration solution can be supercooled to below -70°C and vitrifies without crystallization at approximately -115°C (Sakai et al., 1990).
Ultrarapid cooling by liquid nitrogen
The dehydrated shoot tips were transferred onto sterilized aluminum foil in droplet form using a disposable plastic pipette. Each foil strip held 10 to 20 shoot tips, which were rapidly frozen in liquid nitrogen at -196°C using forceps and stored for a minimum of 1 hour (Fig. 3d,e).
Thawing and recovery procedure
For thawing and unloading, the aluminum foil strip was immersed for 20 minutes in Rewarming Solution (RS), composed of liquid MS medium supplemented with 205.2 g/L (0.6 M) sucrose (㏗: 5.8) (Fig. 3f). To ensure uniform and rapid thawing of the shoot tips, the conical tube was gently shaken at room temperature. The thawed shoot tips were blotted on filter paper to remove any residual RS and then placed onto the CIP potato recovery medium. The recovery medium contained 25 g/L sucrose, 20 mL/L coconut water (C5915, Merck Sigma-Aldrich®), 0.1 ㎎/L gibberellic acid (G7645, Merck Sigma-Aldrich®), 0.4 ㎎/L kinetin (K0753, Merck Sigma-Aldrich®), and 2.8 g/L Phytagel, with the ㏗ adjusted to 5.6.
Assessment of plant viability and recovery
For each cryopreservation attempt, 60 out of 90–100 shoot tips were thawed after at least 1 hour in liquid nitrogen to evaluate initial viability (at 4 weeks) and recovery rate (at 10 weeks) using CIP Recovery medium. The experiment was conducted in triplicate, with 20 shoot tips per replicate. Remaining shoot tips were temporarily stored in cryovials and transferred to the liquid nitrogen tank once a regeneration rate above 40% was confirmed.
During the first stage of recovery, plantlets were incubated in CIP recovery medium under dark conditions at 24°C for two weeks. After this period (Fig. 3g), surviving meristems initiated shoot regeneration with temporary etiolation due to the lack of photosynthesis. The plantlets were then transferred to standard MS medium for continued normal development.
Starting from two weeks post-thaw culture, explants were subcultured every two weeks under standard light conditions (16-hour photoperiod, 100–140 μmol/㎡/s) and a temperature of 24°C ± 1°C. Survival was assessed at week 4, with explants considered viable if they exhibited green spots or callus-like structures with regenerative potential. By week 8, recovery was confirmed only for explants that developed functional organs, such as leaves, stems, or roots (Fig. 3h,i).
Statistical analysis
The cryopreservation experiment was conducted with three replicates, each consisting of a group of 20 shoot tips, resulting in a total sample size (n) of 60 per treatment. For the control group (-LN), 10 shoot tips were used per replicate, with three replicates in total. The control group underwent all cryoprotective procedures except LN freezing. Mean survival and recovery rates for both -LN and +LN treatments are presented as percentages with standard error (SE). Data were analyzed using ANOVA, followed by Duncan’s multiple range test (p < 0.05). Statistical analyses were performed with SAS 7.1 software, and results are reported as mean percentages with SE.
Results and Discussion
This study utilized CIP’s droplet-vitrification protocol to cryopreserve in vitro plantlets from eight potato cultivars developed by Korea’s Highland Agriculture Research Institute (HARI). For cultivars with comparatively low post-thaw regeneration rates, the protocol was optimized with cultivar-specific adjustments to enhance recovery and ensure preservation success. These RDA-bred cultivars feature high anthocyanin content, resistance to environmental stresses and diseases, and excellent processing qualities (Table 1).
Recovery rate in routine cryopreservation
The initial application of the standard CIP protocol evaluated the survival rate (SR) at 4 weeks and the recovery rate (RR) at 8 weeks for each cultivar. The cryoprotective treatment group before liquid nitrogen freezing (-LN, Control) showed high recovery rates of 80–100% at 8 weeks, with no statistically significant differences observed (ANOVA). In the post-thaw recovery group (+LN), assessed based on the development of leaves, stems, and roots, the recovery rates varied by cultivar. ‘Jayoung’ showed the lowest recovery rate at 43.3%, while ‘Arirang 2ho’ exhibited the highest at ‘76%’. Among the eight tested cultivars, ‘Arirang-1ho (71%)’, Arirang 2ho (76%)’, ‘Mangang (71%)’, and ‘Goldenball (68%)’ demonstrated the highest regeneration rates, with RR ranging from 68% to 76%. In contrast, ‘Hongyoung’ showed slightly lower recovery rates (61%), and the lowest regrowth rates were observed in ‘Jayoung (43%)’, ‘Dami (46%)’, and ‘Haryeong (48%)’ (Table 2). The overall plant recovery rate across the eight cultivars was 60.9%. This result falls slightly below the average recovery rate of 65.5% reported in CIP’s large-scale cryopreservation study conducted in 2022 (Vollmer et al., 2022).
Table 2.
Viability response of 8 potato cultivars to CIP’s standard cryo-protocols (CIP, 2024).
Protocol optimization based on initial cryopreservation results
The first round of cryopreservation data highlighted the need for protocol optimization for specific cultivars. Among the eight tested varieties, four cultivars—‘Jayoung (43.3%),’ ‘Dami (46.6%),’ ‘Haryeong (48.3%),’ and ‘Hongyoung (61.6%)’ —exhibited significantly lower regeneration rates, prompting genotype-specific protocol customization. This outcome reflects genotype-specific characteristics, prompting further research on various factors, including material quality and type, cold acclimation during pretreatment, osmoprotective conditions, and mitigation of osmotic stress (Zhang et al., 2023).
The utilization of larger apical shoot tip dimensions
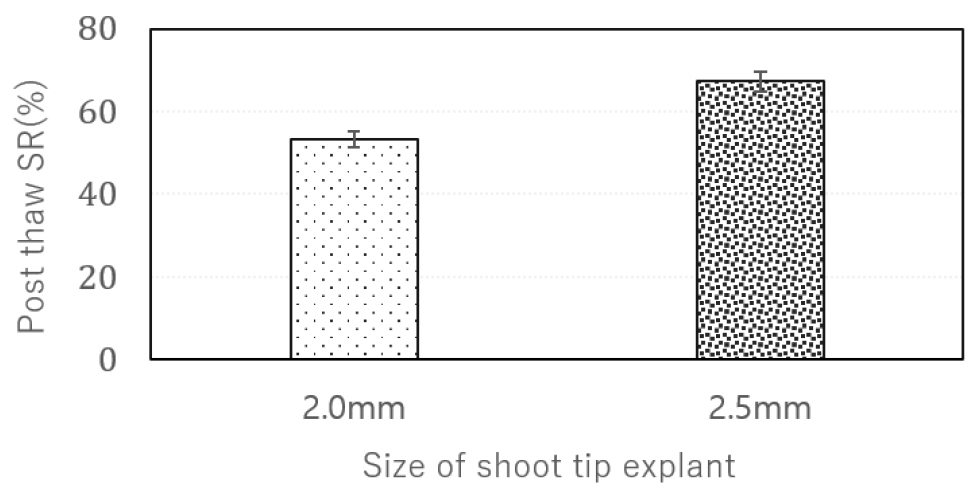
One of the primary adjustments involved increasing the size of the excised shoot tips from 1.5–2.0 ㎜ to 2.0–2.5 ㎜ (Fig. 2). According to the findings of Vollmer et al. (2019), cryopreservation experiments accounted for genotype-specific regeneration responses by varying the age of in vitro-grown plantlets used in the experiments, ranging from 2 to 7 weeks, and adjusting the size of the excised shoot tips from 1 to 4 ㎜ (Vollmer et al., 2019). In the case of the ‘Haryeong’ cultivar, securing a healthy, sterile donor plant proved challenging due to the presence of potential endophytes during in vitro multiplication. The extended period required to eliminate contamination, along with the suboptimal growth condition of the donor plants, negatively impacted the regeneration outcomes. When explants of the same size as those from other cultivars were used, ‘Haryeong’ showed a y lower regeneration rate of 48%. This finding underscored the importance of tailoring explant size and protocol parameters to the specific physiological condition of each cultivar to improve post-thaw recovery. Following the increase in explant size, the survival rate (SR) improved significantly from 53% to 67%, while the recovery rate at 8 weeks has yet to be assessed (N/A, Fig. 4). Meanwhile, larger shoot tips (2–4 ㎜) may require additional modifications, such as extended dehydration and unloading periods, as the unloaded cryoprotectant and incoming water must travel greater distances compared to smaller shoot tips (1–2 ㎜) (Vollmer et al., 2019). Two cryopreservation studies reported by the NAC in 2006 highlighted the need for genotype-specific adjustments by varying the incubation period of donor plants and the size of shoot tip explants for the ‘Degima’ and ‘STN13’ cultivars (Kim et al., 2006; Yoon et al., 2006).
Duration of dehydration
In the first cryopreservation attempts, the ‘Hongyoung’ cultivar exhibited lower survival rates in the -LN treatment group and reduced regrowth rates in the +LN treatment group. To mitigate damage caused by the dehydration solution (PVS 2) and ensure sufficient dehydration to achieve vitrification status, five different dehydration durations, ranging from 30 to 70 minutes in 10-minute increments, were tested. Among these treatments, the standard protocol duration of 50 minutes yielded the highest eight-week regrowth rate of 61% (Fig. 5).
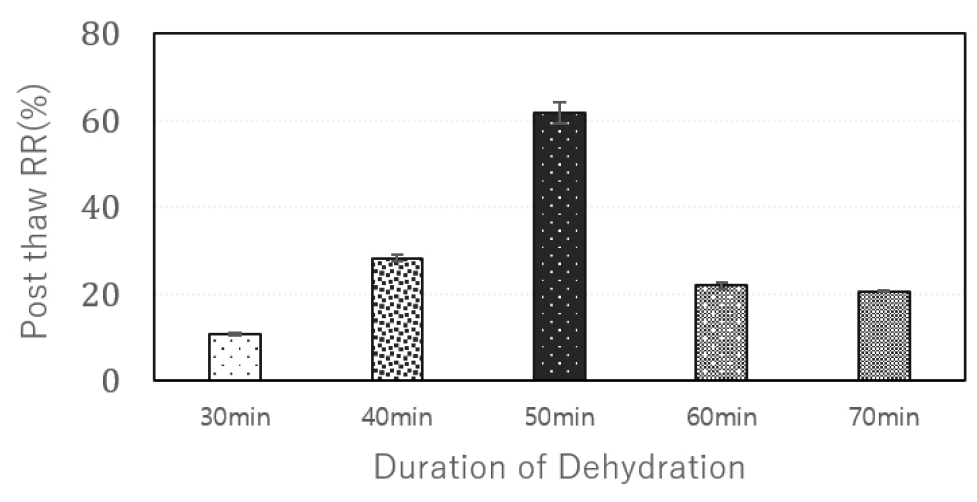
As the dehydration duration deviated from the 50-minute benchmark—either shortened or extended—the regrowth rate declined significantly by week 8 post-thaw. These results suggest that a dehydration duration of 30 to 40 minutes may not be sufficient to achieve adequate tissue dehydration and vitrification in ‘Hongyoung.’ Conversely, extending the dehydration beyond 50 minutes likely exacerbates tissue damage caused by the CPA (PVS2), resulting in diminished regrowth potential. These results are consistent with a previous study, which demonstrated the highest recovery rates in LN-exposed samples after 40–50 minutes of dehydration, with a marked decline in viability observed at 60 minutes (Panta et al., 2014). This underscores the critical need to optimize dehydration time to achieve sufficient tissue dehydration while minimizing CPA-induced damage, particularly given the significant toxicity of PVS2, a highly concentrated vitrification solution, to plant tissues. Sensitivity to dehydration time varies greatly between species, with exposure times ranging from 20 minutes for persimmon (Matsumoto et al., 2001) to 120 minutes for Prunus shoot tips (Niino et al., 1997).
Variation in cold acclimation procedure
The video material available on the CIP website (https://cipotato.org/genebankcip/process/cryopreservation/) provides detailed instructions on a two-phase cold acclimatization process as part of the physiological preparation for cryoprotection. During the final stage of the propagation cycle, the plantlets first undergo mild chilling at 7°C for 10–20 days, followed by a secondary acclimatization phase at 4.5°C for 1–4 days. This two-step cold acclimatization protocol represents a recent improvement not included in Vollmer’s 2022 study.
When the plantlets were cultured in a cold room with low light at 4°C, plant growth was observed to be almost entirely halted. Moreover, certain cultivars, such as ‘Dami,’ exhibited deterioration in plant condition during the 4°C acclimatization stage. In response to this observation, we revised the protocol by applying only the primary cold hardening phase at 7°C before proceeding to the cryoprotection treatment.
This modification resulted in a significant improvement in post-thaw regeneration rates. Plantlets subjected only to the 7°C phase demonstrated an significant increase of over 24% in the eight-week recovery rate compared to those exposed to both phases of the cold acclimatization protocol (Fig. 6). Recent studies highlight the importance of adjusting cold acclimation periods to align with genotype-specific growth patterns, typically ranging from 2 to 3 weeks at 7 ± 2°C with a 16-hour photoperiod and light intensity of 10–20 µmol·m-2·s-1 (Vollmer et al., 2022). Folgado et al. (2015) reported that cold hardening and preculture phases significantly influence the intracellular water content of potato shoot tips, affecting their cryopreservation outcomes. Additionally, studies have shown that cold hardening at 6°C for three weeks significantly enhances post-thaw recovery in drought and frost-tolerant cultivars, confirming that cold treatment plays a crucial role in supporting physiological processes related to stress tolerance (Folgado et al., 2015). Conversely, the same treatment may have adverse effects on cold-sensitive cultivars, potentially causing damage. In such cases, achieving the physiological state required for cryoprotection may be more effectively facilitated through a chemical preculture rather than an extended cold acclimation period. As an example, in the absence of cold acclimation, Yi et al. (2012) reported that, the pre-culture process is a crucial step for the successful cryopreservation of potato.
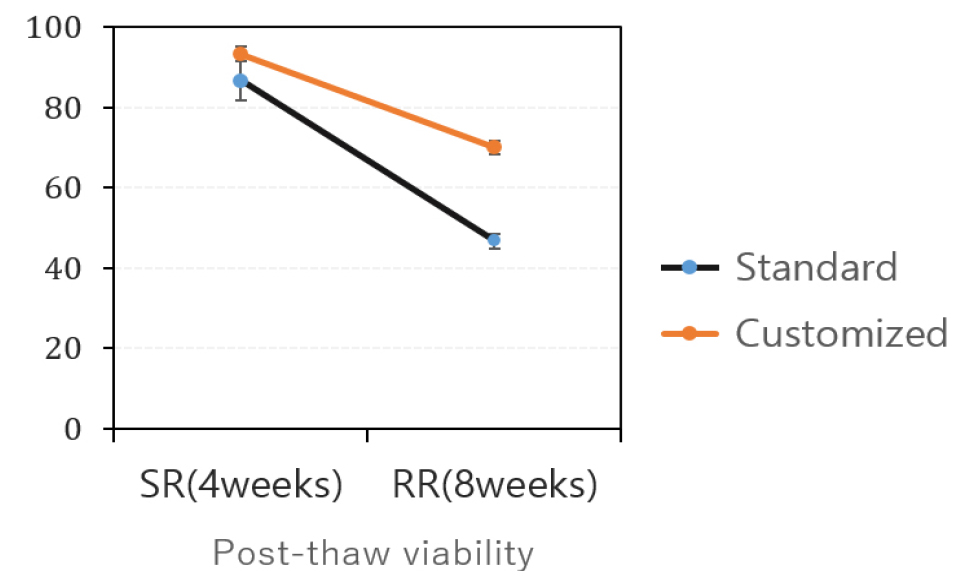
Carbon source adjustment for donor plant optimization
In the tissue culture cycle for mass propagation of samples intended for cryopreservation, certain cultivars exhibited differences in growth rate and morphology based on the sucrose concentration in the culture medium. For example, the ‘Jayoung’ cultivar displayed elongated, slender, and soft stems when grown on a medium containing 20 g/L sucrose. In contrast, plantlets grown on a 30 g/L sucrose medium exhibited stiffer, lignified growth with reduced vigor and a slower growth rate.
When shoot tips were excised from plantlets cultivated under these two media conditions and subjected to cryopreservation, those from the 20 g/L sucrose medium showed enhanced regeneration. Specifically, explants excised from softer donor plants yielded a 30% increase in recovery rate, rising from 43% to 73% (Fig. 7). These findings align with previous reports linking the growth conditions of donor plants to their regeneration capacity. Notably, a study by Panta et al. (2014) demonstrated that potato plantlets grown under blue LED light produced smaller shoot tips, yet exhibited significantly improved post-thaw regeneration rates.
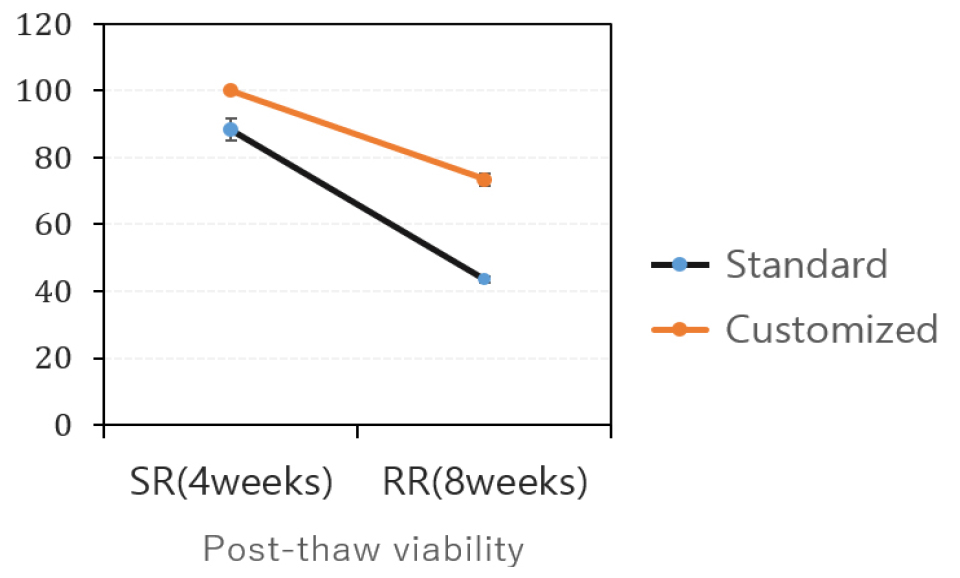
In addition to the sucrose concentration, other studies have explored methods to enhance donor plant vigor. For instance, Lee et al. (2024) reported that overlaying a liquid layer on solid media induced more vigorous growth, resulting in a substantial increase in post-cryopreservation regeneration rates, with improvements of up to 23% compared to plantlets grown solely on solid media. These results emphasize the importance of optimizing culture conditions for donor plants to maximize regeneration potential following cryopreservation.
This study successfully optimized and implemented CIP’s droplet vitrification-based cryopreservation protocol for a range of Korean potato genotypes. As a result, shoot tips exhibiting high regeneration rates have been securely preserved in liquid nitrogen, ensuring long-term conservation for future use. All cryo-preserved accessions data are cataloged within the Germplasm Management System, managed by the National Agrobiodiversity Center, Rural Development Administration (RDA) (https://genebank.rda.go.kr/).
To ensure sustainable preservation of potato genetic resources—one of the world’s major staple crops—an integrated conservation strategy combining field storage, in vitro collections, and cryopreservation within an active, base, and safety (A-B-S) framework is essential. The National Agrobiodiversity Center and Highland Agriculture Research Institute of RDA will prioritize cryopreserving domestic varieties and vulnerable resources, with plans to build a comprehensive cryo-collection by 2026.
