

Introduction
Materials and Methods
Plant material
Apical meristem culture and virus removal treatment
Isolation of RNA and RT-PCR
Data analysis
Result and Discussion
Virus detection of fire blight resistant apple rootstocks G11 and G30 (RT-PCR)
Virus detection of in vitro plants derived from apical meristem culture
Antiviral treatment of in vitro plants derived from apical meristem culture
Comparison of thermotherapy treatment periods for in vitro plants derived from apical meristem culture
Introduction
Apple, a leading fruit crop in South Korea and worldwide, faces significant threat from fire blight. First reported in a Korean apple orchard in Anseong, Gyeonggi-do, in 2015, this disease, caused by Erwinia amylovora (Burill), has since spread nationwide, inflicting serious economic losses on apple and pear orchards (Ham et al., 2020; Lee et al., 2022). Fire blight is a severe disease affecting the Rosaceae family, it can devastate entire orchards (Lee et al., 2018). Most Korean apple orchards use rootstocks such as M.9, which are highly susceptible to fire blight, necessitating an urgent switch to fire flight resistant rootstocks. Consequently, rootstocks such as Geneva (G11, G30, etc.) developed by Cornell University’s Geneva Research Institute and Budagovsky (B9) developed in Russia, known for their fire blight resistance, are being considered.
Apples are primarily propagated through grafting, and virus infections can lead to reduced fruit yields and quality (Cembali et al., 2003). Therefore, distributing disease-free seedlings widely is crucial in mitigating the damages. Common viruses affecting apples include Apple chlorotic leaf spot virus (ACLSV), Apple mosaic virus (ApMV), Apple stem pitting virus (ASPV), and Apple stem grooving virus (ASGV), while viroids such as Apple scar skin viroid (ASSVd) have also been reported (Hu et al., 2019; Keshavarz and Hajnajari, 2019; Li et al., 2020; Xing et al., 2018). Cho et al. (2024) reported on the fruit characteristics of virus-free and virus-infected ‘Hongro’ apple trees. In 2021, during the third year after planting, the average fruit set per virus-free tree was 71.8. In comparison, virus-infected trees had 33.2 fruits (ASGV), 48.0 fruits (ASPV+ASGV), and 68.2 fruits (ACLSV+ASPV+ASGV), representing a reduction in fruit set of up to 54.0% compared to virus-free trees.
Methods for producing virus-free seedlings typically involve several steps. Initially, viruses are eradicated from infected plants through meticulous in vitro culture and heat treatment, followed by precise shoot tip culture (Dĩaz-Barrita et al., 2008; Paprstein et al., 2008). Alternatively, an effective approach employs heat treatment, followed by excision of the apical meristem and vigorous shoot proliferation ex vitro (Vivek and Modgil, 2018). Chemical interventions employing potent antiviral agents such as ribavirin (Hansen and Lane, 1985), 6-azauracil (Hansen, 1988), and 2-thiouracil (Maliogka et al., 2009) on culture media during in vitro plant proliferation are also prevalent. Moreover, hybrid methods incorporating heat treatment with antiviral treatments (Hu et al., 2015) or heat treatment coupled with cryopreservation (Li et al., 2016) have shown enhanced efficacy in virus elimination.
This study aimed to assess the efficacy of various treatments, including heat treatment (37°C), antiviral treatment with ribavirin, and shoot tip culture, either individually or in combination, on in vitro plantlets for the mass production of virus-free seedlings of Geneva rootstocks (G11 and G30). Additionally, the study sought to compare the rates of virus removal using reverse transcription polymerase chain reaction (RT-PCR). Considering the rising occurrence of fire blight in Korean orchards (Myung et al., 2016; Park et al., 2016), investigating virus removal methods for the production of stable virus-free seedlings of fire blight resistant apple rootstocks such as G11 and G30 is imperative. The findings of this research could be of significant value for the future production of virus-free seedlings in fruit tree cultivation, particularly for apples.
Materials and Methods
Plant material
In this experiment, 3-year-old G11 and G30 apple rootstocks, known for their resistance to fire blight, were used. These rootstocks were grafted onto test packaging provided by the Chungcheongbuk-do Agricultural Research and Extension Services. Twenty individuals infected with each of the five viruses (ACLSV, ASPV, ASGV, ApMV, and ASSVd) were selected from each rootstock for virus removal treatments. The presence of the viruses was assessed by RT-PCR to determine the rate of virus-free propagation.
Apical meristem culture and virus removal treatment
Using plantlets of fire blight resistant G11 and G30 apple rootstocks, virus removal tests were conducted through heat treatment, antiviral treatment, and shoot tip culture. For shoot tip culture, the shoot tips were washed in clean water for approximately 30 min, followed by immersion in 70% ethanol for approximately 30 s for surface sterilization. After rinsing three times with sterile water, the shoot tips were subjected to secondary surface sterilization by immersion in a 2% sodium hypochlorite solution for 20 min. Following another round of rinsing, the meristems were dissected under a stereomicroscope (0.1–0.3 ㎜) and cultured on in vitro media for 8 weeks. The culture medium comprised MS (Murashige and Skoog, 1962) basal salts (4.4 g/L), myo-inositol (0.1 g/L), ascorbic acid (0.1 g/L), 6-benzylaminopurine (2.0 ㎎/L), naphthalene acetic acid (0.05 ㎎/L), sucrose (30.0 g/L), and agar (8.0 g/L). Subsequently, 50 mL of media was dispensed in 250 mL culture bottles, autoclaved at 121°C and 1.2 atm for 20 min, and maintained at 23°C ± 1°C under a 16-h light/8-h dark photoperiod (cool white LED, 30 µmol·m-2·s-1). During the culture period, shoot development was observed, and virus testing was performed using leaves formed after 8 weeks of in vitro culture.
Heat treatment involved exposing 20 in vitro plantlets derived from shoot tips to temperatures of 37°C and a relative humidity of 65% for varying durations (0, 10, 20, and 30 days) in a plant growth chamber (VS-8407-1300, Vision, Korea). Antiviral treatment consisted of culturing in vitro plantlets with ribavirin (1-B-D-ribofuransyl-1,2,4-triazole carboxamide, MBcell, USA) at concentrations of 0, 20, 30, and 40 ㎎/L in the culture media at 23°C ± 1°C for 4 weeks.
Isolation of RNA and RT-PCR
Total RNA was extracted from 100 ㎎ of G11 and G30 plant samples using the Hybrid-RTM kit (GeneAll, Korea), and the RNA concentration was quantified using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA). PCR was performed using cDNA as a template with virus-specific primer sets (Table 1).
Table 1.
Primer sequences of apple dwarfing rootstock viruses by RT-PCR analysis.
| List of PCR primers | Product size (bp) | |
| Primersz | Primers sequence (5’-3’) | |
| ACLSV-Fy | GCAGACCCTTCATGGAAAGA | 509 |
| ACLSV-R | CGCAAAGATCAGTCGTAACAGA | |
| ASPV-F | AAGCATGTCTGGAACCTCATG | 368 |
| ASPV-R | GATCAACTTTACTAAAAAGCATAAGT | |
| ASGV-F | GCCACTTCTAGGCAGAAACTCTTTGAA | 273 |
| ASGV-R | AACCCCTTTTTGTCCTTCAGTATACGAA | |
| ApMV-F | CTCCAAACACAACTTTTGATGACTT | 113 |
| ApMV-R | GTAACTCACTCGTTATCACGTACAA | |
| ASSVd-F | ACGAAGGCCGGTGAGAAG | 202 |
| ASSVd-R | CCGCTGCGTCAAAGAAAAAG | |
| nad5-Fx | GATGCTTCTTGGGGCTTCTTGTT | 158 |
| nad5-R | CTCCAGTCACCAACATTGGCATAA | |
The extracted RNA was subjected to PCR using the NBPA2202 P˚CHEK™ Virus/Viroid Detection Kit (Nexbio, Korea) and a thermocycler (C1000 Touch, Bio-Rad Laboratories, USA). The thermal cycling conditions were as follows: 50°C for 30 minutes, 95°C for 15 minutes, followed by 35 cycles of 95°C for 30 seconds, 58°C for 30 seconds, and 75°C for 1 minute, with a final extension at 72°C for 5 minutes. The PCR products were separated by electrophoresis on a 2% agarose gel, with Nad5 (158 bp) used as the control. The following bands were detected: ApMV (113 bp), ASSVd (202 bp), ASGV (264 bp), ASPV (368 bp), and ACLSV (509 bp). Electrophoresis was also performed using the Fragment Analyzer (Advanced Analytical Technologies, USA), and the results were analyzed with the PRO Size 3.0 program to determine the presence of viral infection.
Data analysis
All analytical experiments were repeated twice. In each experiment a set of 20 plants were used for determination of each parameter. Data from each experiment were subjected to Duncan’s multiple range test using SAS program (Version 6.21, SAS Institute Inc., Cary, NC, USA). Further, we conducted Fisher’s LSD test to determine the mean separation. A probability level of 0.05 (p < 0.05) was considered for statistical significance in all data analyses.
Result and Discussion
Virus detection of fire blight resistant apple rootstocks G11 and G30 (RT-PCR)
In this study, plant materials sourced from fire blight resistant apple rootstocks G11 and G30, cultivated at the Chungcheongbuk-do Agricultural Research and Extension Services, were used. Virus testing conducted via RT-PCR detection of the apple rootstocks revealed the predominant coinfection of ACLSV, ASPV, ASGV, ApMV, and ASSVd viruses. The detection of five viruses conducted in G11 and G30 apple rootstocks by RT-PCR. The G11 rootstocks of were infected with ACLSV and ASPV viruses, while the G30 rootstocks were uninfected (Fig. 1).
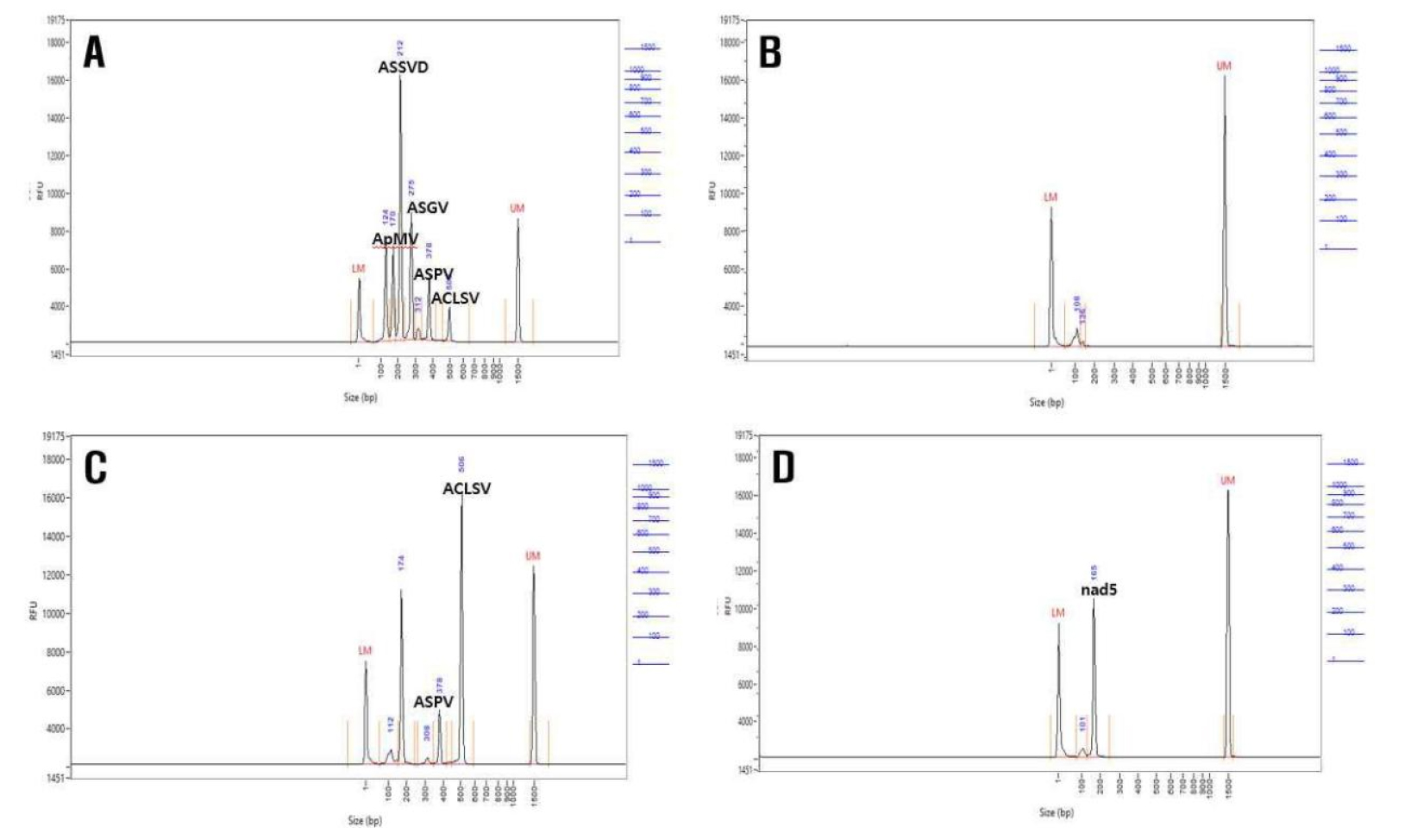
Among ten samples tested for the major five viruses, G11 rootstock showed mixed infections of 2–4 viruses, with ACLSV at 70%, ASPV at 60%, ASGV at 80%, ApMV at 100%, and ASSVd at 20%, whereas G30 rootstock exhibited ASPV at 90%, ASGV at 10%, ApMV at 100%, ACLSV at 60%, and ASSVd at 40% (Table 2). Consequently, to procure samples suitable for virus-free propagation through heat treatment, antiviral treatment, and shoot tip culture, virus-infected samples were specifically targeted for testing.
Table 2.
Detection of viruses G11 and G30 fire blight resistant apple rootstocks using RT-PCR.
| Apple rootstocks |
Number of samplesz | Nad5y | Virus | ||||
| ACLSVx | ASPV | ASGV | ApMV | ASSVd | |||
| G11 | 1 | + | + | + | + | + | − |
| 2 | + | − | + | − | + | − | |
| 3 | + | − | − | + | + | − | |
| 4 | + | + | + | + | + | − | |
| 5 | + | + | + | + | + | − | |
| 6 | + | + | + | + | + | + | |
| 7 | + | + | + | + | + | − | |
| 8 | + | − | − | + | + | − | |
| 9 | + | + | − | − | + | + | |
| 10 | + | + | − | + | + | − | |
| G30 | 1 | + | + | + | − | + | − |
| 2 | + | + | − | − | + | + | |
| 3 | + | − | + | − | + | + | |
| 4 | + | + | + | − | + | + | |
| 5 | + | + | + | − | + | − | |
| 6 | + | − | + | − | + | − | |
| 7 | + | − | + | + | + | − | |
| 8 | + | + | + | − | + | + | |
| 9 | + | + | + | − | + | − | |
| 10 | + | − | + | − | + | − | |
Virus detection of in vitro plants derived from apical meristem culture
Survival rates and virus-free rates of G11 and G30 rootstocks subjected to virus inactivation treatments via RT-PCR method are presented in Table 3. Meristems, excised at sizes ranging from 0.1 to 0.3 ㎜ for culturing, showed virus-free rates for each virus as follows (Fig. 2): 45% for ACLSV, 50% for ASPV, 40% for ASGV, 25% for ApMV, and 95% for ASSVd in G11 rootstock. In G30 rootstock, the virus-free rates were 80% for ACLSV, 25% for ASPV, 35% for ASGV, 30% for ApMV, and 75% for ASSVd. These findings are consistent with those of Plopa and Preda (2013), who documented infection rates ranging from 50% to 58% when employing meristem cultures of apple trees infected with the ApMV. At a meristem size of 0.3 ㎜, the infection rates were 50%–58%, whereas they increased to 77%–85% when the meristem size was 1.0 ㎜. Meristem culture, a prevalent method for virus elimination in various plant species, typically involves culturing meristematic tissues of 0.2–0.4 ㎜, proving effective in virus removal (Faccioli and Marani, 1998). Momma and Takahashi (1983) found that meristem cultures of hop plants infected with hop stunt viroid effectively removed the viroid when using meristems of sizes between 0.2 and 0.3 ㎜ but not when using meristems >0.4 ㎜, indicating a decrease in virus removal efficiency with larger meristem sizes. While meristematic regions are typically expected to be devoid of viruses (Grout, 1999), plant RNA viruses spread systemically through the plant’s vasculature, similar to the movement of plant metabolites (Leisner et al., 1993). Cho et al. (2016) noted that the extent of viral infection is influenced by the distribution of viruses within stem cross-sections and by their spread to meristems. Consequently, achieving virus-free apple seedlings through meristem culture alone presents a challenge because of the inability to completely deactivate viruses using single treatment methods.
Table 3.
RT-PCR results of virus detection measurements after shoot apical meristem culture from fire blight resistant G11 and G30 apple rootstocks.
|
Apple rootstock |
Number of samplesz |
Virus-free rate of plants (%) | Virus-free plants by virus type (%) | ||||
| ACLSVy | ASPV | ASGV | ApMV | ASSVd | |||
| G11 | 20 | 30 | 45 | 50 | 40 | 25 | 95 |
| G30 | 20 | 20 | 80 | 25 | 35 | 30 | 75 |

Fig. 2.
Production process of virus-free fire blight resistant G11 and G30 apple rootstocks in vitro shoot apical meristem culture. (A) Axillary bud, (B) Apical meristem culture with a size of 0.2 mm, (C) In vitro shoot derived from apical meristem culture, (D) In vitro culture of multiple shoots from plants.
Antiviral treatment of in vitro plants derived from apical meristem culture
In vitro plantlets from shoot tips of G11 and G30 rootstocks treated with ribavirin at concentrations of 0, 10, 20, and 30 ㎎/L for 4 weeks. The survival rate of in vitro plants decreased as the ribavirin concentration increased, while the virus-free rate gradually increased (Table 4). In G11 rootstock, treatment with ribavirin 20 ㎎/L resulted in a 100% survival rate with a 30% virus-free rate. However, treatment with ribavirin 30 ㎎/L and 40 ㎎/L decreased the survival rate from 88.3% to 72.0%, with observed leaf deformation symptoms indicating abnormal plant growth (Table 4, Fig. 3). Similarly, in G30 rootstock, ribavirin 20 ㎎/L treatment showed a 100% survival rate with a 51.0% virus-free rate. Treatment with ribavirin ≥30 ㎎/L resulted in survival rates of 80.3%–88.3% and virus-free rates of 70.3%–85.0%; however, normal plant characteristics were not observed, which indicated ribavirin 20 ㎎/L as the most appropriate concentration.
Table 4.
Effect of in vitro ribavirin treatment on rate of survival and virus-free plants and virus-specific free plant in G11 and G30 apple rootstocks by RT-PCR after 4 weeks.
|
Apple rootstock |
Ribavirin (㎎/L) |
Survival rate (%) |
Virus-free rate of plant (%)z | Virus-free plants by virus type (%) | ||||
| ACLSVy | ASPV | ASGV | ApMV | ASSVd | ||||
| G11 | 0 | 100aX | 15.0d | 15.0c | 25.3b | 80.0a | 25.3c | 95.3a |
| 20 | 100a | 30.3c | 70.3b | 75.0a | 90.7a | 30.3bc | 100a | |
| 30 | 88.3b | 45.3b | 95.3a | 54.7ab | 95.3a | 45.3b | 100a | |
| 40 | 72.0c | 60.3a | 95.3b | 71.3a | 95.3a | 65.7a | 100a | |
| G30 | 0 | 100a | 20.0c | 85.0a | 20.0b | 70.3b | 29.7c | 100a |
| 20 | 100a | 51.0b | 76.0a | 65.3a | 94.3a | 51.0bc | 100a | |
| 30 | 88.3b | 70.3ab | 89.7a | 80.0a | 100a | 70.3ab | 100a | |
| 40 | 80.3c | 85.0a | 94.3a | 85.0a | 100a | 85.0a | 100a | |
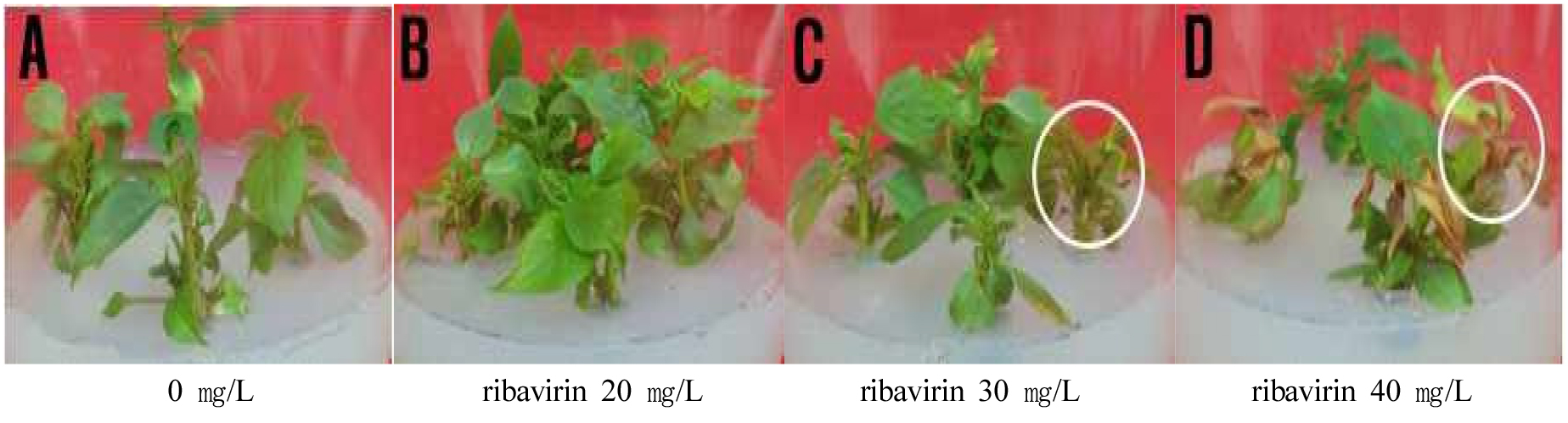
Fig. 3.
Production process of virus-free apple rootstock plants using antiviral treatment. (A) Control, (B) In vitro normal plants treated with antiviral (ribavirin 20 ㎎/L) in medium, (C) In vitro abnormal plants treated with antiviral (ribavirin 30 ㎎/L) in medium, (D) In vitro abnormal plants treated with antiviral (ribavirin 40 ㎎/L) in medium.
Antiviral agents are effective in removing viruses; however, high concentrations may induce phytotoxicity and weaken plant vigor. Major symptoms of antiviral treatment include chlorophyll degradation and necrosis of leaf margins (El-Dougdoug et al., 2010). Use of the antiviral agent ribavirin in culture medium to produce virus-free apple seedlings effectively removed the ACLSV (Hansen and Lane, 1985). Ma et al. (2024) reported that the eradication of apple ASGV and ACLSV were increased with thermotherapy duration (36℃ for 16 hours in light and 32℃ for 8 hours in darkness), reaching over 60% after two weeks and 100% after four weeks, regardless of salicylic acid treatment.
However, after 8 weeks of treatment, the survival rate sharply decreased to 30.0% in the group treated with ribavirin 20 ㎎/L, despite showing 100% survival rates after 2 and 4 weeks. Moreover, survival rates gradually decreased to 20.0% with ribavirin 40 ㎎/L treatment over time, indicating that the optimal duration of antiviral treatment is up to 4 weeks (Choet al., 2016).
Comparison of thermotherapy treatment periods for in vitro plants derived from apical meristem culture
During the thermotherapy period at 37°C, in vitro plants derived from apical meristem culture showed varying virus removal rates (Fig. 4). For G11 rootstock, subjecting the plantlets to 10 days of high temperature treatment resulted in comparable virus removal rates, with a virus-free rate ranging from 24.0% to 30.3%. Extending the treatment to 20 days increased the survival rate to 89.6% with a virus-free rate of 61.0%. However, after 30 days, although the virus-free rate reached 74.7%, the survival rate dropped to 20.0%, making it challenging to obtain normal individuals. In contrast, G30 rootstock exhibited a 100% survival rate and a 35% virus-free rate after 10 days of treatment. However, after 20 days, the survival rate decreased to 30.3%, whereas the virus-free rate remained at 100% (Table 5). Chun et al. (2022) noted that higher shoot growth rates during high-temperature treatment increase the likelihood of obtaining virus-free meristems. They also observed that high temperatures restrict virus replication and movement (Cooper and Walkey, 1978; Zhao et al., 2018).

Table 5.
Effect of thermotherapy treatment at 37℃ on rate of survival and virus-free plant during in vitro culture in G11 and G30 apple rootstocks.
| Apple rootstock |
Period of thermotherapy in days (37°C) | Number of samplesz |
Survival ratey (%) |
Virus-free rate of plantsx (%) |
| G11 | 0 | 20 | 100a | 24.0b |
| 10 | 20 | 100a | 30.3b | |
| 20 | 20 | 89.6a | 61.0a | |
| 30 | 20 | 20.0b | 74.7a | |
| G30 | 0 | 20 | 100a | 24c |
| 10 | 20 | 100a | 35b | |
| 20 | 20 | 30.3b | 100a | |
| 30 | 20 | 00.c | nd |
The decline in survival rates during high-temperature treatment may be attributed to the inhibitory effect of higher temperatures on shoot differentiation and growth, despite effectively inhibiting virus movement and replication (Hu et al., 2015; Ozgur et al., 2004). Thus, an optimal temperature is necessary to produce virus-free plantlets by inhibiting virus activity while minimizing adverse effects on plant growth. Wang et al. (2006) reported that in vitro cultured pear shoot tips were subjected to 37°C for 25, 30, and 35 days, the highest virus elimination efficiency was achieved by combining thermotherapy for 35 days with the culture of 1.0 ㎜ meristem tips.
These findings align with previous studies on virus removal efficiency in in vitro plantlets of new apple cultivars following high-temperature treatment and meristem culture. Lee et al. (2013) reported similar trends, with 84% of ACLSV removed from 31 individuals, 68% of ASGV removed from 31 individuals, and 83% of ASSVd removed from 24 individuals.
Furthermore, the vigorous growth of plantlets during high-temperature treatment in apples facilitated the production of virus-free plantlets, emphasizing the importance of prioritizing plant growth rate over virus activity (Hollings, 1965).
The results obtained from treating in vitro plantlets derived from meristem culture of G11 and G30 cultivars with ribavirin at 20 ㎎/L followed by 20 days of high-temperature treatment at 37°C revealed promising outcomes. For the G11 cultivar, a high survival rate of 95% was achieved, with notable virus removal rates for different virus types: ACLSV 85%, ASPV 90%, ASGV 70%, ApMV 100%, and ASSVd 100%, resulting in an overall virus-free rate of 70%. However, the G30 cultivar exhibited exceptional results with a 100% survival rate and a 100% virus-free rate. The challenges encountered in virus removal for the G30 cultivar, attributed to multiple virus infections, were evident compared to single infections (Knapp et al., 1995; Paprstein et al., 2008).
When comparing individual virus removal methods, meristem culture emerged as the most effective, with virus-free rates of 31.7% for the M26 cultivar with high-temperature treatment, 21.7% with chemical treatment, and 50.0% with meristem culture (Kwon et al., 2019). Additionally, Kim et al. (2017) reported successful ACLSV virus removal at 100% when culturing various calluses ranging from 0.4 ㎜ to 1.2 ㎜ derived from dormant buds of the “Hongro” apple tree. Most instances resulted in approximately 93.5% virus removal for ASSVd, except for when culturing meristematic and lateral callus tissues separately. This study underscores the need for efficient micrografting techniques facilitating grafting of buds smaller than 1.0 ㎝ under conditions where shoots grow at temperatures >36°C to enhance virus-free rates through high-temperature treatment (Chun et al., 2022).
The research findings advocate for the simultaneous application of all three-methods meristem culture, ribavirin treatment at 20 ㎎/L, and high-temperature treatment at 37°C for 20 days as a more effective approach than individual treatments. Hence, it is recommended to use ribavirin at a concentration of 20 ㎎/L, ensuring minimal toxicity to in vitro plantlets derived from meristem culture, for producing virus-free plantlets of G11 and G30 cultivars, coupled with high-temperature treatment at 37°C for 20 days for optimal outcomes.
