

서 언
재료 및 방법
조사지역 및 입지환경조사
출현식물, 식생조사 및 분석
토양분석
결과 및 고찰
입지환경 분석
군집구조
군집별 출현식물
상대우점치 분석
종다양성지수 분석
유사도지수 분석
토양 분석
Ordination 분석
적 요
서 언
최근 전통지식을 기반으로 한약재로서 이용해오던 약용자원식물은 천연원료에 관한 관심이 집중되고, 경제적 가치가 증대됨에 따라 바이오산업의 소재로써 재평가되고 있으며(Park et al., 2012), 최근에는 이러한 약용식물을 활용해 천연물신약, 기능성식품 및 화장품의 원료로서 광범위하게 활용되고 있다(Song et al., 2020). 또한 생물다양성협약(CBD), IUCN Redlist, Cites 등 세계적으로 희귀한 식물을 비롯한 자생식물에 관한 관심이 높아지는 동향으로(Falk and Olwell, 1992; Lee et al., 2011) 자생식물의 보전을 넘어서 연구기반을 활용한 특허권 경쟁으로 이어지고 있다.
그 중 피자식물문(Angiospermae), 산토끼꽃목(Dipsacales), 인동과(Caprifoliaceae), 인동속(Lonicera) 식물은 적응력이 뛰어난 경우가 많다. 대표적으로 우리나라의 자생식물이자 미국의 침입외래식물인 괴불나무(Lonicera maackii)는 미국에서 자생종을 위협하고 있으며(David and Donald, 2003), 인동(Lonicera japonica)의 경우 유럽, 뉴질랜드 등지에서는 침입외래식물로 지정되어 있을 정도로 그 번식력과 적응력이 뛰어나다(Kristina, 2010). 동시에 중국에서 인동은 금은화라고 하여 향균, 항염증, 항당뇨 등 다양한 효능이 입증돼 차, 술, 음료 등에서 활용되고 있으며(Ye et al., 2018), 괴불나무 또한 약용자원으로서의 잠재적 가치가 크다고 한다(An et al., 2018).
연구의 대상종인 길마가지나무(Lonicera harae)는 인동과 인동속의 낙엽관목으로 자생지는 중국 동북부, 일본의 쓰시마섬, 한국에서는 황해도 이남의 산지에 분포하며, 주로 남부 지역을 중심으로 자생하고 있다(Kim and Kim, 2011). 형태는 특산식물인 숫명다래나무(Lonicera coreana)와 유사한 형태적 특성을 지닌 다른 종으로 구분되어 왔다(Ishidoya and Jeong, 1923). 이후 인동속에 관한 연구에서도 두 종의 꽃, 화분, 열매에 분류학적 차이는 없으나 소지, 엽병 등에 강모의 발생여부에서 차이를 보였으며, DNA 다형성 분석에서 차이점을 보이고 있다고 했다(Jeon, 1994). 하지만 최근에는 형태학적으로 작은 차이를 두고 별개의 종으로 구분하지 않는 경향을 보이고 있다(Kim and Kim, 2011; Korea National Arboretum, 2021).
관련 연구로는 형태학적 연구(Jeon, 1994), 발아특성(Ko et al., 2018) 정도로 수행되었을 뿐, 자생지 및 생육환경에 대한 연구는 매우 미흡한 실정이다. 이와 달리 길마가지나무는 봄을 알리는 전령사로, 꽃의 향취가 좋아 천연향료자원으로서의 쓰임새가 있을 뿐 아니라 Chung et al. (2017b)에 따르면 “꼬리볼레”라는 이름으로 식용하기도 하여 잠재적인 가치를 지니고 있다.
따라서 본 연구는 우리나라에 자생하는 길마가지나무의 생육환경특성 및 입지환경 조사를 통해 자생지를 보존 및 복원하고 향후 산업화 가치가 있는 천연물 소재의 대량생산 기술개발에 기초자료로 활용하고자 한다.
재료 및 방법
조사지역 및 입지환경조사
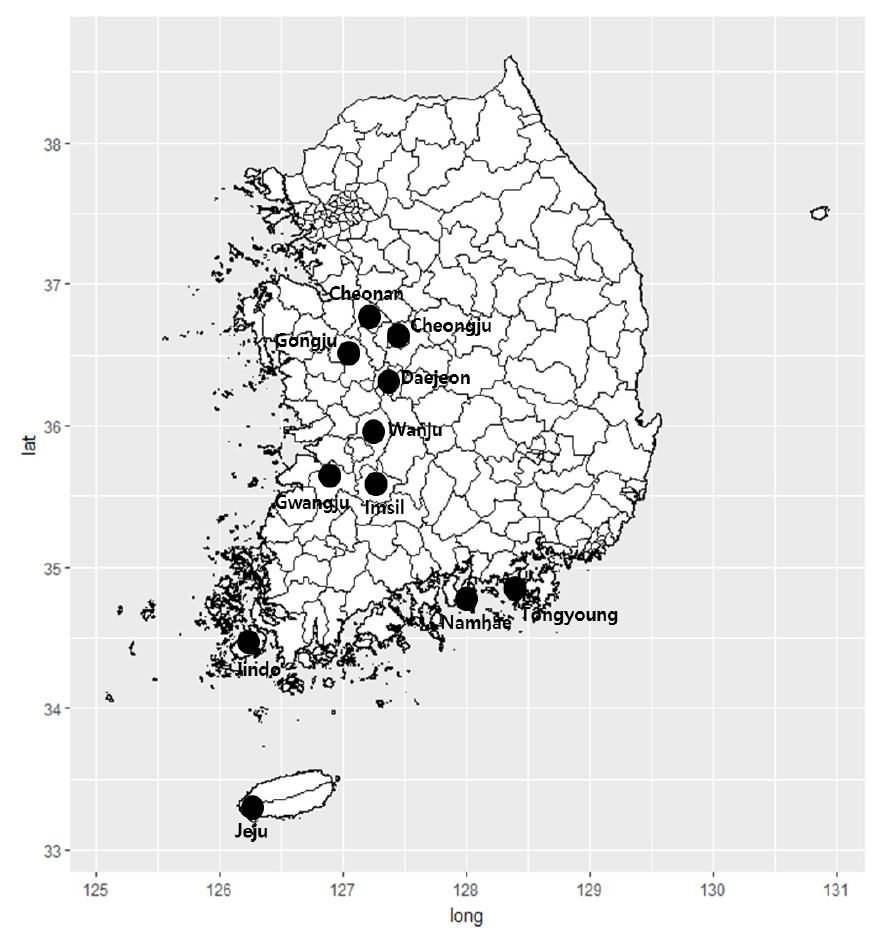
연구대상지역은 문헌자료를 바탕으로 길마가지나무 자생지를 파악하였으며, 현장조사를 통해 선정하였다. 자생지는 충청도 공주, 청주, 천안 3개 지역, 전라도 광주, 진도, 완주, 임실 4개 지역, 경상도 대전, 남해, 통영 3개 지역, 제주도로 종합 11개 지역 24개 방형구(10 × 10 m)를 선정하였다(Fig. 1). 자생지 입지환경은 GPS (Garmin, GPSMAP64S)와 Sunnto Clino Compass를 이용하여 위도・경도, 고도, 경사 및 방위를 측정하였으며, 기상환경은 기상청의 통계연보를 참고하였으며, 최근 10년의 연평균온도, 최고기온평균, 최저기온평균, 연평균강수량, 상대습도, 최소상대습도를 참고하였다.
출현식물, 식생조사 및 분석
자생지의 출현식물 및 식생조사는 2020년 3월부터 2020년 11월까지 약 9개월간 길마가지나무 및 출현식물을 쉽게 확인할 수 있는 개화시기, 결실기를 중심으로 실시하였다. 출현식물은 표본 채취 및 현지에서 야장에 기입하였다. 식물의 분류는 Engler의 분류체계(Melchior, 1964)에 준하였으며, 학명 및 국명은 국가표준식물목록(Korea National Arboretum, 2017)에 준하여 작성하여 목록을 정리하였다(Appendix 1). 식물의 동정은 Lee (2003a, 2003b; Kim and Kim, 2011)의 식물도감을, 한국특산식물목록(Chung et al., 2017a)을, 침입외래식물은 Korea National Arboretum (2016)을 참고하여 정리하였다. 채집한 표본 및 화상자료는 국립산림과학원 산림약용자원연구소 표본실에 보관하였다.
조사된 방형구 24개소의 식생조사 방법은 Braun-Blanquet의 방법(Braun-Blanquet, 1964)을 따라 실시하였으며, 출현 식물은 교목층(8 m 이상), 아교목층(2 ~ 8 m), 관목층(0.8 ~ 2 m), 초본층(0.8 m 이하)으로 수고별로 구분하였다. 수고별로 구분된 층은 식피율과 개체수를 기록하여 층위별로 Curtis and McIntosh (1951)의 중요치(I.V.: Importance Value)를 백분율로 나타낸 상대우점치(I.P.: Importance Percentage)를 산출하였고, 수관의 층위별 가중치를 부여하여 평균상대우점치 (M.I.P.: Mean Importance Percentage)를 도출하였다. 층위별 종다양성, 균일성은 Shannon-Weaver의 종다양도(H')를 적용하여 최대종다양도(Maximum H')는 H' max = logS (S는 종수), 균등도(J')는 J' = H'/H'max 및 우점도(D)는 D = 1-J'의 식을 이용하였다. 유사도 지수는 Jaccard 계수(1900, 1901, 1908)를 개선한 Sorensen (1948)의 유사도지수 및 상이도지수를 산출하였다. 자생지간 유연관계의 분석은 출현식물의 중요치를 바탕으로 군집분석을 실시하였다. 군집분석은 PC-ORD v. 5.17 (McCune and Mefford, 2006) 다변량 분석 프로그램을 이용해 군집분석을 실시하였으며, 출현빈도가 5% 미만의 종을 제외하여 나타냈으며 지표종분석을 통해 적절한 군락수를 결정하였다. 전체군락은 환경과의 상관관계를 확인하기 위해 비계량형 다차원 척도법(Nonmetric Multidimensional Scaling)을 이용하여 분석하였다.
토양분석
산림토양은 산림에 자생하는 동・식물의 생명을 유지하는데 필요한 물과 양분을 저장하고 공급하는 지지기반이다. 이러한 성질은 생육환경을 알아보기 위한 중요한 역할을 하므로 토양의 이화학성과 물리적인 특성을 조사하였다. 토양은 Litter층을 제거한 10 ~ 20 ㎝의 깊이에서 방형구당 3개의 시료를 채취하였다. 토성은 피펫법(Pipet method)에 의한 미국 농무부분류기준을, 유기물함량(OM)은 Walkley-Black법(Walkley and Black, 1934), 유효인산(P2O5)은 Lancaster법(Ryu et al., 1977), 질소(T-N)는 Kjeldahl법(1883), 치환성양이온은 1N- NH4OAc (pH 7.0) 침출에 의한 I.C.P분석, 양이온치환용량(C.E.C)은 1N-NH4OAc (pH 7.0) 침출에 의한 NH4-N 킬달 증류법으로 산출하였다. 전기전도도와 pH는 채취된 시료를 1:5로 희석하여 EC meter (HANNA, HI 98331)와 pH meter (HANNA, HI 99121)로 측정하여 평균치를 산출하였다.
결과 및 고찰
입지환경 분석
길마가지나무 자생지 11지역 24개소의 입지환경을 조사한 결과(Table 1), 해발고도는 평균 173 m로 가장 높은 자생지는 진도의 첨찰산 483 m, 낮은 곳은 남해의 물건리 해안방조림 8 m로, 산지의 저지대를 중심으로 등산로, 계곡부 및 해안림에 분포하였다. 경사는 평균 8.5°로 가장 경사가 급한 곳으로 대전 만인산이 25°, 완만한 곳으로 남해 물건리 2°로 확인되었다. 대부분 10° 내외의 경사지에서 자생하는 것을 보았을 떄 급경사지에서도 생육이 가능하나 완경사지를 선호하는 것으로 확인됐다. 사면방위는 동남, 남동, 북서, 동, 남, 북향으로 나타나 방위에 큰 영향을 받지는 않으나 대체로 남향에 분포하는 것으로 나타났다. 개체수의 경우 최소 제주도 청수곶자왈 2개체부터 최대 공주 계룡산의 18개체, 평균 8개체가 군락을 이루는 것으로 확인되었다.
Table 1.
Environmental and vegetation characteristics of surveyed plots
자생지 11개 지역의 기상환경을 확인한 결과(Table 2), 연평균기온은 13.7℃로 최저지역은 임실 11.7℃, 최고지역인 제주 15.9℃로 큰 차이를 보이고 있었다. 평균 최고기온은 19.0℃로 최저지역은 임실 18.3℃, 최고지역은 광주 및 남해 19.7℃, 큰 차이를 보이고 있지 않았다. 최저기온 평균은 9.2℃로 최저지역인 임실 6.1℃, 최고지역인 제주 13.4℃로 지역 간 큰 차이를 보이고 있었다. 평균상대습도는 70.2%로 최저지역인 청주 62.0%, 최고지역은 제주 77.0%로 나타나 연평균습도에서는 큰 차이를 보이고 있지 않았다. 평균최소습도는 23.1%로, 최저지역인 임실 및 금산 19%와 최고지역인 제주 37.0%와 큰 차이를 보이고 있었다. 연평균강수량은 1386.5 ㎜로, 최저지역인 진도 1121.2 ㎜와 최고지역인 남해 1839.4 ㎜와 큰 차이를 보이고 있었다.
Table 2.
Basic weather information of surveyed regions (2011 ~ 2020)
군집구조
군집구조 분석은 전체 출현식물을 대상으로 Outlier Analysis를 통해 특이종과 전체 군집중 5%이하 출현종을 제외한 24개 군락 118종을 대상으로 PC-ORD v. 5.10 (McCune and Mefford, 2006)의 DCA분석을 실시하였다. 나누어진 군락은 지표종 분석(Indicator species analysis)을 통해 가장 유의한 3개 군집으로 구분되었다(Fig. 2). 1번 군집은 아까시나무가 우점하는 6개 식생(Community I), 2번 군집은 느티나무가 우점 하는 7개 식생(Community Ⅱ), 3번군집은 일본잎갈나무가 우점 하는 11개 식생으로(Community Ⅲ) 나누어졌다. 군락은 지역별로도 구분이 됐으나 지역 내에서도 구분되는 군락도 확인할 수 있었다(Fig. 3). 아까시나무가 우점하는 1번 군집은 1, 2, 7, 10, 16, 24에 해당하는 대전지역 일부, 천안, 진천, 제주지역 일부가 해당되었으며, 느티나무가 우점 하는 2번 군집은 3, 4, 5, 8, 9, 14, 15에 해당하는 공주, 진천, 남해지역이 해당되었고, 느티나무가 우점 하는 3번 군집은 6, 11, 12, 13, 17, 18, 19, 20, 21, 22, 23에 해당하는 천안, 광주, 진도, 통영, 완주, 대전지역 일부로 가장 많은 군락이 해당되었다. 군집1인 아까시나무 군집은 지표종으로 아까시나무, 사위질빵, 소태나무가, 군집2인 느티나무 군집은 느티나무, 칡, 누리장나무가, 군집3인 일본잎갈나무 군집은 국수나무, 때죽나무, 주름조개풀이 지표종으로 나타났다.
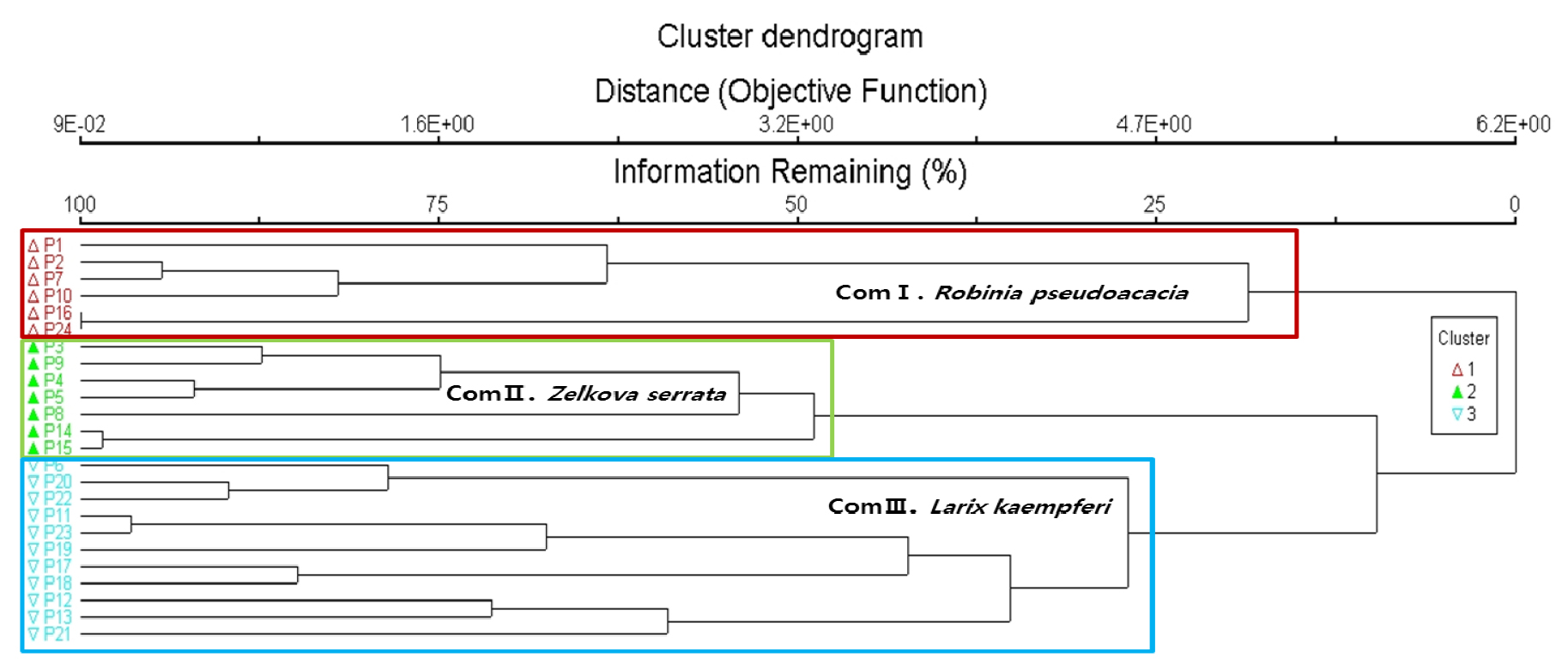
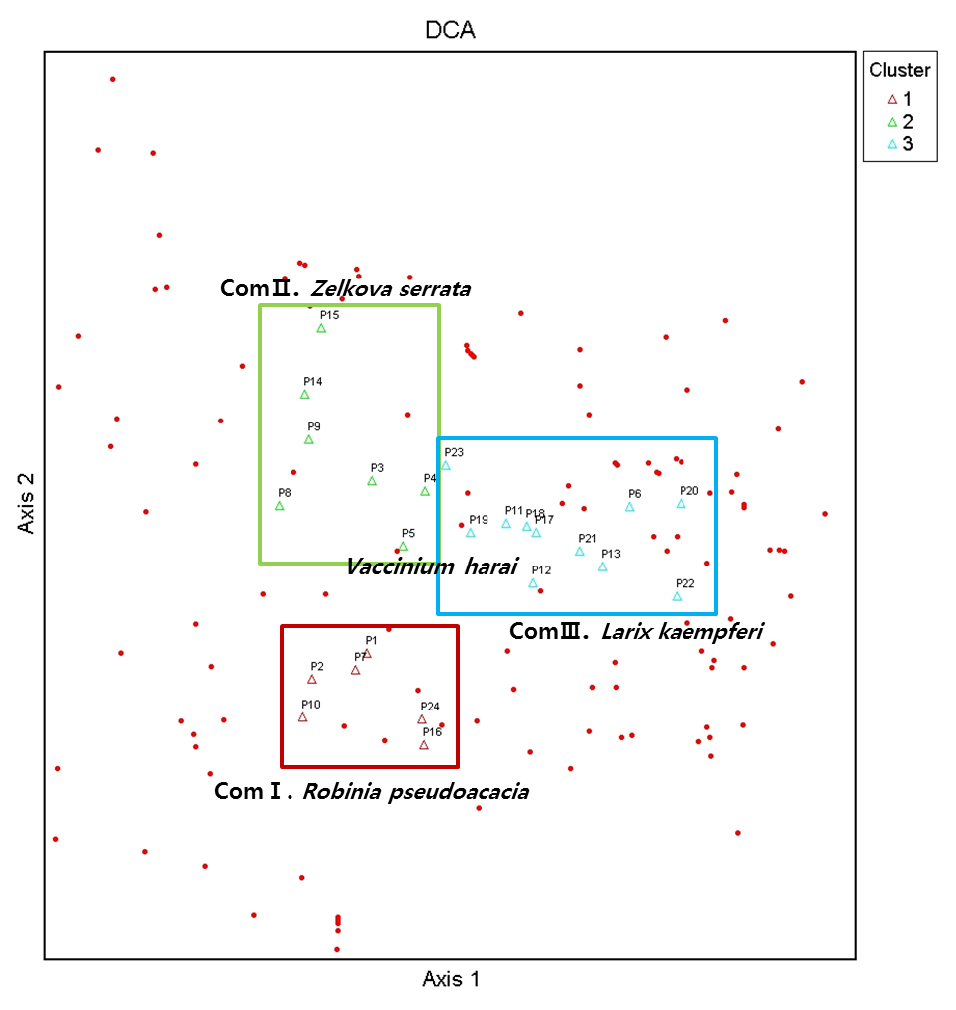
군락전체에서 가장 출현이 많은 종은 느티나무, 쥐똥나무, 주름조개풀, 으름덩굴, 사위질빵 등으로 산림저지대의 임연부, 등산로 주변에서 흔히 확인할 수 있는 종이 대표적인 지표종으로 나타났다.
군집별 출현식물
자생지에서 출현한 관속식물은 80과 166속 198종 3아종 24변종 4품종 총 229분류군으로 확인되었다(Table 3). 이 중 특산식물 병꽃나무 1분류군이 확인되었으며, 침입외래식물 미국자리공, 족제비싸리, 아까시나무 등 5과 9속 9종 총 9분류군, 산림청지정희귀식물로 LC등급에 해당하는 녹나무, 쥐방울덩굴, 이팝나무 등 4과 5속 5종 총 5분류군으로 나타나, 길마가지나무 자생지의 출현식물은 입지환경 특성상 특이식물이 상대적으로(Byeon et al., 2020) 매우 적은 것으로 판단된다.
Table 3.
Summary on the flora of study area
군집별로는 아까시나무가 지표종인 군집 I에서 출현한 식물은 50과 77속 78종 2아종 11변종 총 91분류군으로, 지표종인 기존의 군락에 침입하여 우점한 것으로 보여 지며, 느티나무가 지표종인 군집Ⅱ에서 출현한 식물은 51과 82속 80종 2아종 10변종 1품종 총 93분류군으로, 11개로 가장 많은 방형구가 해당되며 일본잎갈나무가 지표종인 군집Ⅲ에서 출현한 식물은 61과 111속 129종 2아종 13변종 3품종 총 147분류군으로 가장 많은 분류군을 확인할 수 있었다.
상대우점치 분석
자생지의 군집 및 층위별 상대우점치(I.P.)를 확인한 결과(Table 4), 군집I(아까시나무 군집)의 교목층은 아까시나무(50.98%)가 상층을 우점하고 있었으며, 아교목층은 아까시나무(18.76%)와 종가시나무(14.88%)가 경쟁관계에 있었다. 관목층은 길마가지나무(13.52%)와 쥐똥나무(13.58)가 경쟁관계에 있었으며, 초본층은 으름덩굴(9.31%), 주름조개풀(8.97%)이 경쟁관계에 있었다.
Table 4.
Importance percentage (IP) and mean importance percentage (MIP) of major species in each plant communities
군집Ⅱ(느티나무 군집)의 경우 교목층에서는 느티나무(36.43%)가 우점 하였으며 갈참나무(20.99%), 밤나무(14.29%)가 경쟁관계에 있었다. 아교목층은 느티나무(31.49%)가 우점하고 있었으며, 관목층은 길마가지나무(15.09%)와 신나무(16.69%)가 경쟁관계를 이루고 있었다.
군집Ⅲ(일본잎갈나무 군집)은 교목층에서 일본잎갈나무(20.00%)와 상수리나무(15.51%)가 경쟁관계에 있었으나 특정종이 우세한 모습을 보이지 않았다. 아교목층 또한 때죽나무(13.54%)와 느티나무(12.36%)가 경쟁관계에 놓여있었다. 관목층은 길마가지나무(11.03%)가, 초본층은 주름조개풀(12.88%)이 우점하고 있었다.
군집I은 아까시나무가 상층의 대부분을 구성하는 냉온대 남부 저산지대의 대표적인 식생으로, 장기적으로 길마가지나무의 상층을 이루는 대표적인 지속군락의 한 유형으로 판단된다(Cho and Kim, 2005). 군집Ⅱ는 느티나무가 상층 대부분을 구성하면서 갈참나무와 관목층의 쥐똥나무, 길마가지나무가 층을 이루는 군락이다. 이는 전국에서 확인 할 수 있는 전형적인 느티나무 군락으로(Kong et al., 2012) 길마가지나무와 함께 안정적인 생육환경을 이루고 있음을 확인할 수 있었다. 군집Ⅲ은 일본잎갈나무와 함께 상수리나무, 느티나무와 때죽나무가 경쟁관계에 있는 식생유형을 이루고 있음을 확인할 수 있었다.
종다양성지수 분석
자생지를 구성하는 출현종의 피도를 통해 Shannon의 종다양성지수(H'), 최대종다양성지수(H'max), 균등도(J'), 우점도(D')를 산출한 결과(Table 5), 군집 내에서 군집구조의 복잡성을 나타내는 종다양성지수(H')는 군집Ⅲ이 1.427로 가장 높았으며, 군집Ⅱ가 1.377로 가장 낮은 수치를 보였다. 최대종다양성지수(H'max) 또한 군집Ⅲ이 최대 1.456, 군집Ⅱ 1.413으로 가장 낮은 수치를 보였다. 군집별 종구성의 균일한 정도를 나타내는 균등도(J')는 평균 0.978로 1에 가까운 값을 나타낼수록 종별 피도와 빈도가 균일한 상태를 의미하며(Brower and Zar, 1977), 군집간 편차는 크지 않아 길마가지나무 군집의 종구성이 매우 균일한 것으로 확인되었다. 우점도(D')는 평균 0.022로 한 종이 우점 하는 경향보다는 여러 종이 우점 하는 식생유형을 보이고 있음을 확인할 수 있었다.
Table 5.
Species diversity index of woody and herbaceous species in the investigated plots
유사도지수 분석
유사도지수는 종간, 군집 간 유사도를 통해 서로 비슷한 종이나 생물이 살아가는 공간이 비슷한 곳을 나타내는 지수로서(Lee et al., 2011), 군집 간 20% 미만일 경우에는 서로 이질적인 집단, 80% 이상일 경우 서로 동질적인 집단으로(Whittaker, 1956) 판단할 수 있다.
유사도지수를 산출한 결과(Table 6), 평균 49.10로 나타났으며, 군집I-군집Ⅱ가 55.6%로 가장 유사한 군집으로 확인되었으며, 군집I-군집Ⅲ 49.21%, 군집Ⅱ-군집Ⅲ 42.42% 순 으로 유사도지수가 낮아졌다. 이러한 결과는 길마가지나무의 생육환경이 다양한 환경에서 고유의 군락을 형성하기보다, 군집에 출현한 지표종을 기반으로 다양한 환경의 지역에 적응할 수 있는 특징이 반영된 것으로 판단된다.
Table 6.
Similarity index(%) between communities
| Dissimilarity Index(%) | ||||
| Community IZ | Community IIY | Community IIIX | ||
| Similarity index (%) | Community I1 | 44.34 | 50.79 | |
| Community II1 | 55.66 | 57.58 | ||
| Community III1 | 49.21 | 42.42 | ||
토양 분석
자생지에서 채집한 토양을 분석한 결과(Table 7), 토성은 모래, 미사, 점토의 구성비 평균은 각 56.92%, 19.88%, 18.26%로 모래의 비중이 높은 사양토가 대부분을 차지했으며, 군집I은 제주지역 양질사토, 진천지역 식양토, 군집Ⅱ에서 진천지역 식양토, 군집Ⅲ에서 천안지역 양질사토로 확인되었다. 갈색산림토양이 대부분이었으며 암적색 산림토양이 일부 확인되었다. 토양산도는 평균 pH 4.95로 최저 pH 3.70부터 최대 pH 6.22로 산성토양에 해당했다. 군집별로는 군집I이 평균 pH 4.85로 가장 낮았으며, 군집Ⅱ와 군집Ⅲ의 경우 각 pH 5.24, pH 5.26으로 큰 차이를 보이지 않았다. 유기물 함량은 평균 15.23%로 군집Ⅱ가 23.93%로 가장 높았으며, 군집I 11.22%, 군집Ⅲ 8.18% 순으로 큰 차이를 보였다. 전질소 함량은 평균 0.66%로 군집Ⅱ가 1.01%로 가장 높았으며, 군집I 0.57%, 군집Ⅲ 0.32%순으로 나타났다. 유효인산의 경우에는 평균 196.19 mg kg-1, 군집II 309.11 mg kg-1로 높았으며, 군집Ⅲ 145.88 mg kg-1, 군집I 125.86 mg kg-1순으로 전체적으로 매우 높은 수치를 보이고 있었다. 양이온치환용량(CEC) 또한 평균 32.93 cmolc kg-1으로 군집Ⅱ에서 최대 43.72cmolc kg-1, 군집I 32.43 cmolc kg-1, 군집Ⅲ 20.48 cmolc kg-1순으로 전체적으로 높았다. 치환성양이온의 평균 함량은 각각 Ca2+ 18.95 cmolc kg-1, Mg2+ 3.26 cmolc kg-1, Na+ 0.10 cmolc, kg-1, K+ 0.44 cmolc kg-1 로 확인되었다. 전기전도도(EC)는 평균 0.37 ds/m-1로 나타났다.
토양분석 결과에 따르면, 군집간 차이는 있으나 대체적으로 물리・화학성은 전형적이지 않은 갈색산림토양, 암적색산림토양의 성질을 보이고 있었다. 물리적 성질은 토양배수가 양호한 사양토, 양토가 대부분이었는데 이는 우리나라 평균 산림토양이 모래, 미사, 점토의 구성비가 각각 41.7%, 41.5%, 16.8% (Ministry of Land, Infrastructure and Transport, 2017)와 비교했을 때 모래가 높은 산림토양임을 확인할 수 있었다. pH는 우리나라 산림토양 평균 pH 5.48 (Jeong et al., 2002) 보다 낮은 산성토양이었으나 최대 pH 6.22을 확인한다면 길마가지나무에 어느 정도 토양내성이 있는 것으로 판단된다. 화학성 중 보비력의 지표인 유기물함량, 유효인산, 양이온치환용량이 평균 산림토양(Jeong et al., 2002)에 비해 수치가 높아 양질의 토양에서 생육하고 있는 것으로 판단된다.
Table 7.
Soil characteristic in each vegetation colony of Lonicera harae
Ordination 분석
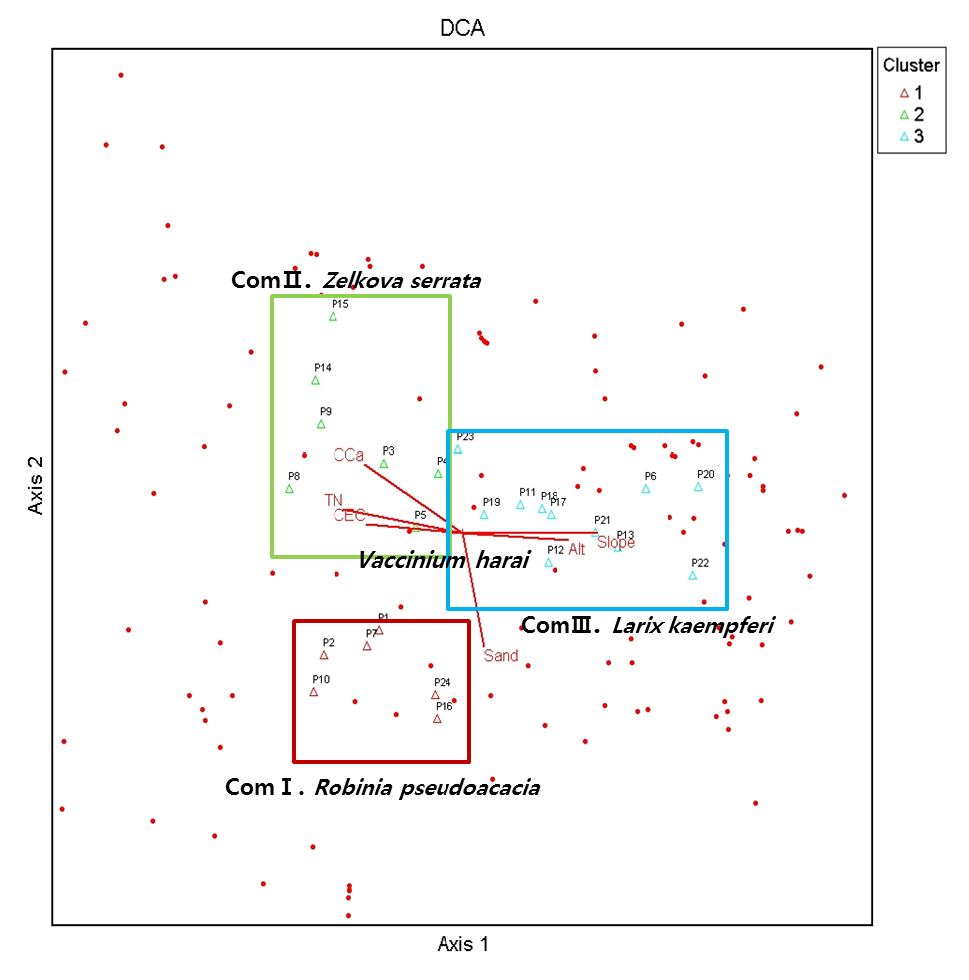
분석된 군집을 바탕으로 군집과 환경 및 토양의 물리・이화학성간 상관관계를 알아볼 수 있는 DCA 분석을 PC-ORD v. 5.10 (McCune and Mefford, 2006) 통해 분석한 결과(Fig. 4), 각 플롯은 길마가지나무를 중심으로 환경적 요인에 의해 나뉘었다고 볼 수 있다. 특히 군집Ⅱ에 해당하는 3, 5, 8, 9, 14 방형구 대부분이 토양의 이화학적 요인인 치환성 Ca2+, 전질소(TN) 함량, 치환성양이온(CEC)의 영향에 따라 분포하고 있음을 확인할 수 있었으며, 군집Ⅲ에 해당하는 6, 11, 12, 13, 17, 18, 19, 20, 21 방형구 대부분이 해발고도(Alt), 경사(Slope)와 같은 환경에 의해 분포하고 있음을 확인할 수 있었다. 군집Ⅰ에 해당하는 1, 2, 7, 10, 16, 24 방형구는 토양의 이화학성과, 물리적 성질인 모래의 비율이 군락 형성의 요인이라고 판단된다.
적 요
길마가지나무의 자생지 11지역 24개소의 입지환경을 조사한 결과 길마가지나무는 주로 산지의 낮은 지대를 중심으로 임연부, 등산로주변, 계곡부 및 해안림 곳곳에 분포하고 있었으며, 경사는 대부분 평지나 완경사지를 중심으로 분포하고 있었다. 사면방위는 가리지 않았으나 주로 햇볕을 많이 받는 남향에 주로 군락을 이루고 있었다. 군집구조는 분석결과 각각 아까시나무(군집Ⅰ), 느티나무(군집Ⅱ), 일본잎갈나무(군집Ⅲ)가 지표종인 3개 군집으로 나뉘었다. 출현식물의 경우 군집 I에서 출현한 식물은 50과 77속 78종 2아종 11변종 총 91분류군, 7개의 방형구로 느티나무가 지표종인 군집Ⅱ에서 출현한 식물은 51과 82속 80종 2아종 10변종 1품종 총 93분류군, 11개로 가장 많은 방형구가 해당되며, 느티나무가 지표종인 군집Ⅲ에서 출현한 식물은 61과 111속 129종 2아종 13변종 3품종 총 147분류군으로 나타났으며, 특이식물로는 특산식물 1분류군, 침입외래식물 9분류군, 희귀식물 5분류군으로 상대적으로 적은 개체수가 출현하였다. 상대우점치(I.P.) 분석결과 각 군집의 지표종으로 대표되는 종이 있으나 특정군집에 독립적으로 출현하는 경우는 적었다. 대표적으로 느티나무, 참나무, 아까시나무, 밤나무 등이 있었으며, 아까시나무군집의 교목층을 제외하면 대부분 경쟁관계에 있음을 확인할 수 있어 지속적인 관찰이 필요하다고 판단된다. 종다양성지수 분석과 유사도지수 분석에서도 상대우점치 분석과 유사한 결과를 확인할 수 있었는데, 군집 간 특정종이 우세하기 보다는 균등도가 매우 높으며, 출현식물이 상이하기 보다는 유사한 경향을 띄고 있었다. 토양분석 결과, 군집 간 전형적인 갈색산림토양, 암적색산림토양의 성질을 보였으며, 배수가 좋은 사양토를 기반으로 하고 있었다. 유기물의 경우에는 일반적인 산림토양에 비해 함량이 대체적으로 높은 경향을 보이고 있었다. 미루어 보았을 때 길마가지나무의 자생지 분포는 지역적인 경향보다는 상층이 울폐하지 않는 임연부, 등산로를 중심으로 유기물이 풍부한 환경에 분포해 환경에 대한 적응력이 뛰어난 것으로 판단된다. 하지만 상층이 울폐한 경우에는 피압으로 인해 생육이 부진한 경향을 보여 광 요구도가 높은 수종으로 판단된다.
