

서 언
주걱댕강나무(Diabelia spathula ta Siebold & Zucc.)는 일본의 특산식물로 알려져 있었으나 2003년 경남 양산시 소재 천성산에서 수 백 그루가 발견되었다(Joongang Ilbo, 2003; National Institute of Biological Resources, 2012; Yonhap News Agency, 2003). 주걱댕강나무는 천성산 일부지역으로 분포범위가 극히 제한되고 있어 IUCN 적색목록 범주 및 기준에 따라 산림청・국립수목원에서는 멸종위기종(Critically Endangered, CR)으로(Korea National Arboretum, 2012), 환경부 국립생물자원관에서는 관심대상종(Least Concern, LC)으로 평가 및 관리되고 있는 희귀식물이다(National Institute of Biological Resources, 2012). 희귀식물(Rare Plants)은 통상적으로 생태계의 건강한 구조와 기능을 지속하기 위한 보전생물학적 가치가 높아 개체군 및 생육 환경 특성과 관련한 보전연구가 필요하다(Jung et al., 2016). 희귀식물의 제한된 분포역과 개체군의 협소, 이에 수반한 자생지의 종 다양성 부족과 유전적 부동(Genetic drift)에 의한 유전다양성의 감소는 생물다양성의 질적 저하로 이어질 우려가 있어 선제적으로 종 보전 관리 방안이 마련되어야 하며(Kim et al., 2011), 최근 생물다양성을 유지할 수 있는 보전전략의 관심이 높아져 현지 내 보전의 중요성이 더욱 강조되고 있다(Yoon et al., 2020).
주걱댕강나무의 생태를 보면 4월 하순부터 5월 중순까지 황백색 또는 미색을 띠는 꽃을 피우는데, 새 가지 끝에 두 개씩 달리는 깔때기 모양의 꽃은 댕강나무속 중에서 가장 크고, 미려하여 자원식물로서 가치 또한 높은 식물이다. 주걱댕강나무의 분포지는 중국 남부・일본 혼슈 이남에 분포하며(Kim and Kim, 2011), 국내에서는 유일한 생육지가 천성산으로 확인된 것 이외에는 지금까지 생육환경이나 분포범위, 식생구조 등 일체의 연구 자료가 전무한 실정이다. 천성산 일대에 대한 관속식물 분포(Han et al., 2010; Shin and Lee, 2009)와 식생(Cho, 1990; Choi et al., 2005; Park, 2005)에 관한 연구에서는 천성산 전역에 걸쳐 참나무류가 우점하고 일부 능선부에 소나무가 우점하는 것으로 보고하였으나 모든 조사에서 주걱댕강나무는 출현하지 않았다. 이에 본 연구는 현지조사 자료와 연구문헌자료를 종합하여 주걱댕강나무가 생육하는 층위별 식생의 구조를 확인하고 주걱댕강나무가 생육하는 다양한 환경인자를 파악하여 대체서식지 조성에 필요한 자료를 확보하고자 하였다. 또한 개체군 동태를 모니터링하여 종의 속성을 이해하고 지속가능성 여부와 다양한 위협요인 등을 파악하여 적절한 보전관리방안을 제시하고자 하였다.
한편 Diabelia속은 Abelia속에서 분계한 바 있으며(Landrein, 2010; Wang et al., 2015), 우리나라의 주걱댕강나무 샘플을 채집하여 유전구조를 분석한 결과, Diabelia ionostachya 종들과 가까운 유연관계를 가진 식물로 보고한 바 있으나(Zhao et al., 2019) 우리나라에서는 최근 학명의 재정립으로 Diabelia spathulata Siebold & Zucc.으로 표기하였다. 따라서 본 연구에서는 국가표준식물목록(http://nature.go.kr/kpni/SubIndex.do)상의 학명(Diabelia spathulata Siebold & Zucc.)으로 표기하였다.
재료 및 방법
대상지 개황
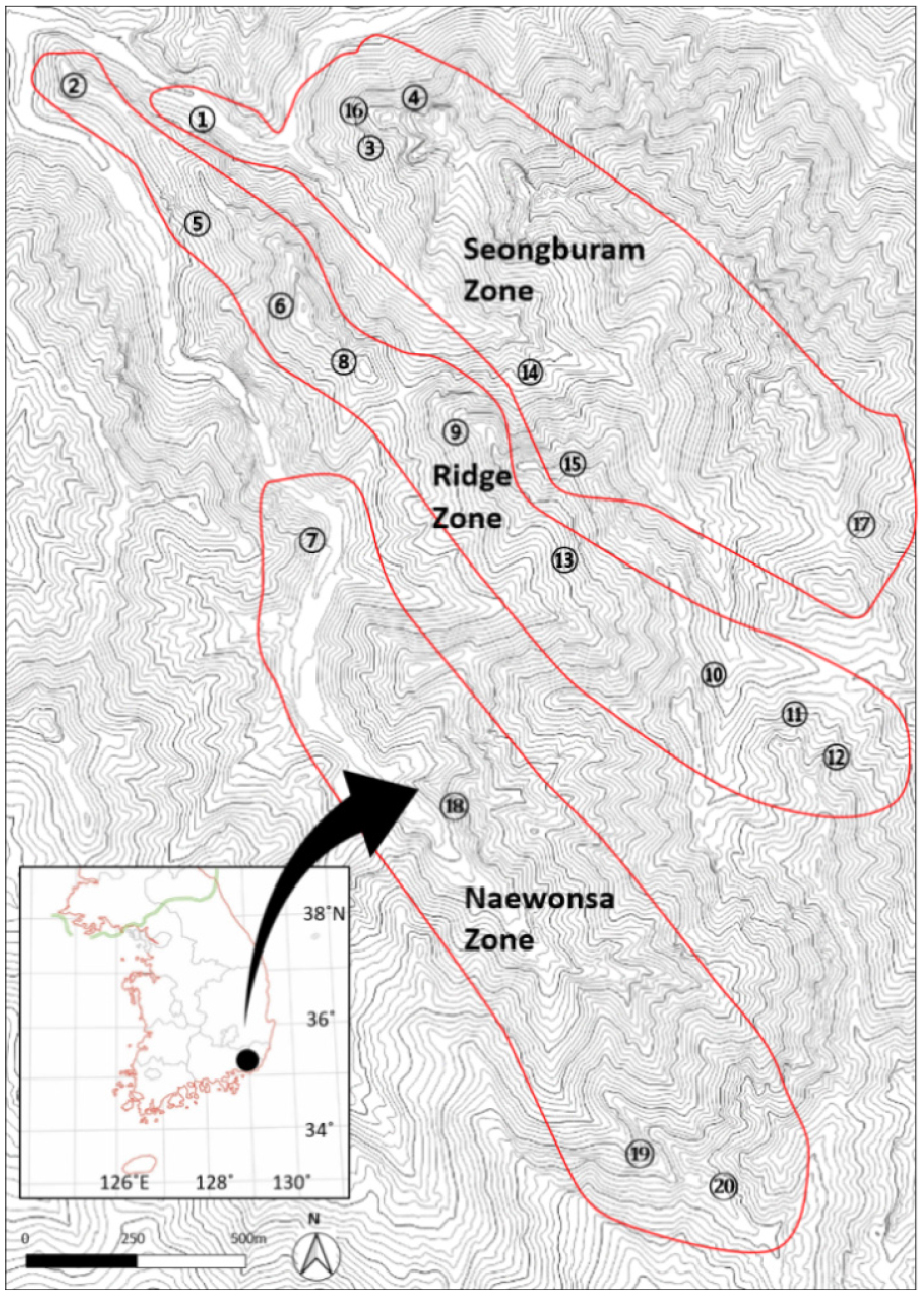
주걱댕강나무의 국내 유일한 생육지로 알려진 천성산(千聖山)은 경상남도 양산시 평산동, 소주동과 상북면・하북면의 경계에 있는 산으로 태백산계에 속하며, 양산시에서 이전의 원효산(元曉山)을 천성산 주봉(922 m)으로, 이전의 천성산을 제2봉(812 m)으로 명칭 변경(1999년)을 하여 오늘에 이르고 있으며, 원효대사가 중국에서 화엄경을 배우러 온 1,000명의 수행자에게 설법을 통해 성인으로 만들었다는 설화가 전해지고 있다. 천성산의 기후대는 남부내륙형 기후에 속하며, 식물구계학적으로 남부아구에 속한다(Han et al., 2010; Shin and Lee, 2009). 천성산과 인접한 기상관측소(기상청 양산 상북, 표준지점 905) 데이터를 분석한 결과, 지난 10년간(2009∼2019년) 연평균 기온은 14.9℃, 평균최고기온 26.25℃, 평균최저기온 10.32℃, 강수량 1,461.03 ㎜, 상대습도 62.28%, 연평균일조량 2,290.89시간으로 나타났다(Korea Meteorological Administration, 2020). 주걱댕강나무의 주 생육지인 천성산 공룡능선과 내원사 일대의 지질과 암질은 산성 화산암류, 유문암질 응회암으로 이루어져 있으며(Kim and Sung 1996; Son et al., 2003), 토양은 급경사 산악지에 분포하는 무등통(Mudeung Series)의 갈색 내지 암갈색 산림토로 미사질이며, 유효토심이 얕고 매우 강산성을 띤다(National Institute of Agricultural Sciences, 2020). 수계는 화북면 용연리 일대 북서향으로 깊은 골짜기가 형성되어 양산천으로 연결되어 있다. 주걱댕강나무의 분포실태를 파악하기 위해 2018년 3월부터 6월까지 예비조사를 통해 주걱댕강나무의 전반적인 생육범위를 확인하였으며, 이번 조사에서 주걱댕강나무 개체군별 해발고도와 사면방향, 그리고 미세지형 내 개체군 형성 정도, 사면별 생육 위치를 감안하여 20개소의 조사구를 선정하였다. 특히 주걱댕강나무 분포지가 햇빛이 어느 정도 확보되는 능선부 또는 사면부에 매우 불연속적으로 출현하고 있어, 격자형 방식 등 일률적인 거리나 고도 적용이 곤란하여 반복적인 답사와 분석을 통한 조사구 위치 조정으로 객관성을 보완하였다. 연구대상지 구획은 천성산 내 익성암 앞 상리천을 따라 형성된 성불암지구(7개소), 상리천과 내원사계곡 사이에 형성된 능선지구(9개소), 내원사계곡을 따라 형성된 내원사지구(4개소) 등 3개 지역으로 나누어 조사를 실시하였다(Fig. 1).
식생구조
식생조사는 종조성 군락규모, 수고, 분포거리, 방위, 고도, 미세지형 등을 감안하여 10 × 10 m (100 ㎡) 크기의 조사구 20개소에서 2019년 5월부터 2020년 4월까지 조사를 실시하였다. 식생조사에 따른 식물동정은 신초발생・개화시기・생육정도 등을 감안하여 계절별로 관찰이 용이한 식물을 분류한 후 반복 답사하여 관찰하였다. 주걱댕강나무 서식처의 식생구조를 파악하기 위해 각 조사구내 출현하는 전 분류군을 대상으로 교목층(8 m 이상), 아교목층(2 ~ 8 m), 관목층(0.8 ~ 2 m), 초본층(0.8 m 이하)으로 구분하여 분류군별 개체의 수고, 수관폭, 흉고직경, 근원직경을 측정하였다. 이를 기반으로 상대밀도(R.D.)와 상대피도(R.C.)를 구한 다음 종별 상대우점치(I.P.={상대밀도+상대피도}/2) 및 평균상대우점치(M.I.P.)를 층위별로 산출하였다(Brower and Zar, 1977; Curtis and Mclntosh, 1951).
군집분류는 조사구별 상대우점치 산출 결과를 반영하여 TWINSPAN에 의한 Classification (Hill, 1979) 분석기법을 적용하여 분류하였으며, 군집간 유사도지수를 통해 유사도(Buell, 1966; Cox, 1976), Shannon-Wiener의 종다양도(H'=-∑pi (logpi), pi는 어떤 종의 개체수 대 전체종의 총 개체수의 비), 최대종다양도(H'max=logS, S는 구성종수), 균재도(J'=H'/H'max), 우점도(D=1-J')를 분석하였다(McNaughton, 1967; Pielou, 1975). 식물의 학명과 국명은 국가표준식물목록(Korea National Arboretum, 2020; Korean Plant Names Index Committee, 2020)에 준하여 작성하였다.
생육 환경
주걱댕강나무 개체군 특성에 관한 데이터를 수집하기 위해 GPS (Oregon300, Garmin)를 이용하여 조사구별 위・경도 및 해발고도를 측정하였으며, 경사계(Tandem-360PC/ 360R G, Suunto)를 이용하여 사면방위와 경사를 측정하여 미세지형을 파악하였다.
토양
토양의 온도와 습도 측정은 토양수분계(EC-300, AQUATERR)를 이용하였으며, 각 조사구별 일기조건을 맞추기 위해 2019년 10월 15일부터 17일까지 3일간에 걸쳐 09:00 ~ 11:00 시간대에 일괄 측정하였다. 토양의 이화학적 특성을 파악하기 위해 각 조사구에서 표토 유기물층을 걷어낸 후 깊이 0 ~ 10 ㎝에서 토양을 500 ㎎씩 채취한 뒤 음건하여 유기물함량(O.M), 전질소(T.N), 유효인산(A.P), 치환성양이온(K+, Ca2+, Mg2+, Na+), 토양산도(pH) 및 전기전도도(E.C.), 양이온치환용량(C.E.C.)을 경상북도농업기술원에 의뢰하여 분석하였다.
빛 환경
주걱댕강나무 개체군 내 빛 환경 특성을 파악하기 위해 디지털카메라(MarkⅡ 5D, Canon)와 어안렌즈(EF 8 ~ 15 ㎜ f/4L Fisheye USM Lens, Canon)를 이용하였다. 조사구별 주걱댕강나무의 수고를 감안 1 ~ 1.8 m 높이에서 하늘을 향한 수직방향으로 천공사진을 5회 촬영하였다. 촬영한 화상자료는 Gap Light Analzer Version 2.0 프로그램을 이용하여 흑색(수관)과 백색(천공 하늘)으로 이원화된 영상을 통해 수관열림도(Canopy openness)를 산출하였다(Fig. 2). 조사구별 측정값의 오류를 최소화하기 위해 산출 시스템에 촬영지점 좌표, 해발고도 및 방위, 경사 등 미세지형 측정값을 더해 보정을 하였다.

엽록소 함량
주걱댕강나무 엽록소(chlorophyll) 함량은 2019년 8월 17일부터 19일까지 엽록소계측기(SPAD-502, Minolta)를 이용, 조사구별로 수고(H: 2.2 m)가 유사한 개체 중 개화한 주걱댕강나무 5개체씩을 선정하여 각 개체당 가지 상부의 성엽 10장씩 3반복 측정으로 평균값을 산정한 후 수관열림도와 상관성을 분석하였다.
개체군 특성
주걱댕강나무 개체군 구조와 개화특성을 파악하기 위해 조사구별 주걱댕강나무의 모든 개체수 및 개체군의 수고를 측정하였으며, 이를 밀도로 환산하였다. 개화특성을 파악하기 위해 2019년 5월 3일부터 6일까지 개화일에 맞추어 모든 개체군의 개화여부와 개화 시 개체의 수고를 확인하여 개화율과 개화특성을 파악하였다.
상관분석
이와 같은 현장조사 결과를 토대로 조사구별 지형 및 토양조건, 수관열림도 등 환경적인 요인과 주걱댕강나무의 개체밀도, 개화율과의 상관성을 분석하였고, 수관층위별 주걱댕강나무의 생육에 따른 타 수종의 영향성을 검토하였다.
결과 및 고찰
대상지 개황
주걱댕강나무 개체군의 범위는 북위 35°24' 58"에서 35°26' 35" 동경 129°05' 43"에서 129°07' 04" 사이였으며, 해발고도는 98∼592 m에서 능선과 사면, 계곡부 등 사방에 불연속적으로 개체군을 형성하고 있음을 확인하였다(Table 1). 각 조사구에서 측정된 전 분류군의 평균 수고는 교목층 8 ~ 13 m, 아교목층 3.4 ~ 6 m, 관목층 1.2 ~ 1.7 m, 초본층 0.2 ~ 0.7 m 범위였으며, 층위별 평균 근원직경은 교목층 15 ~ 31 ㎝, 아교목층 4 ~ 12 ㎝, 관목층 1 ~ 2.2 ㎝, 초본층 0.3 ~ 0.5 ㎝였다(Table 1).
Table 1.
Characteristics and vegetation status of studied plots
식생구조
식물군집의 구조
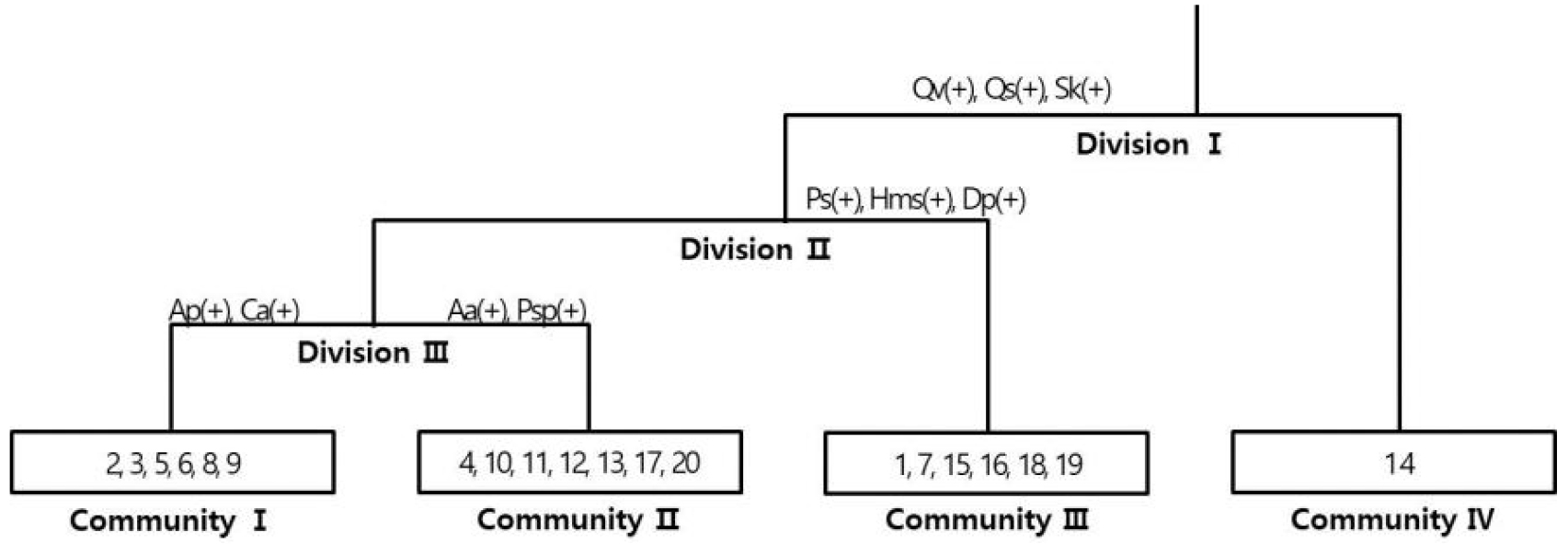
20개 조사구에 출현한 식물을 토대로 TWINSPAN (Hill, 1979)에 의한 classification 분석을 실시한 결과 4개의 군집으로 분류되었다(Fig. 3). 제1단계 Division Ⅰ은 굴참나무(Qv: Quercus variabilis Blume), 졸참나무(Qs: Quercus serrata Murray), 노각나무(Sk: Stewartia koreana Nakai ex Rehder) 유무에 따라 구분되었으며, 제2단계 Division Ⅱ는 굴피나무(Ps: Platycarya strobilacea Siebold & Zucc.), 산수국(Hms: Hydrangea macrophylla (Thunb.) Ser. subsp. serrata (Thunb.) Makino), 꼬리말발도리(Dp: Deutzia paniculata Nakai) 유무에 의해 구분되었다. 제3단계 Division Ⅲ에서는 팥배나무(Aa: Aria alnifolia (Siebold & Zucc.) Decne.)와 잔털벚나무(Psp: Prunus serrulata Lindl. var. pubescens (Makino) Nakai), 당단풍나무(Ap: Acer pseudosieboldianum (Pax) Kom.)와 밤나무(Cc: Castanea crenata Siebold & Zucc.)가 식별종(Indicator species)이었다. 그룹Ⅰ은 조사구 2, 3, 5, 6, 8, 9, 그룹 Ⅱ는 조사구 4, 10, 11, 12, 13, 17, 20, 그룹 Ⅲ은 1, 7, 15, 16, 18, 19, 그룹 Ⅳ는 14로 구분되었다(Fig. 3).
상대우점치
군집별 상대우점치(I.P.)를 분석한 결과(Table 2), 군집 Ⅰ (소나무 군집)은 교목층에서 소나무(I.P.: 78.1%)와 신갈나무(I.P.: 16.8%)가 함께 출현하였으나, 소나무의 세력이 높은 우세를 보였다. 아교목층은 신갈나무(I.P.: 26.7%), 쇠물푸레나무(I.P.: 23.3%), 소나무(I.P.: 15.3%)가 주요 출현종이었고, 관목층은 주걱댕강나무(I.P.: 45.8%)가 우점하는 가운데 털진달래(I.P.: 27.4%), 철쭉(I.P.: 12.7%), 쇠물푸레나무(I.P.: 9.2%)가 함께 출현하였다. 초본층에서도 주걱댕강나무(I.P.: 54.4%)가 우점종이었으며, 쇠물푸레나무(I.P.: 9.2%), 철쭉(I.P.: 7.4%), 조릿대(I.P.: 4.8%), 털진달래(I.P.: 3.2%)가 더불어 출현하였다(Table 2). 군집 Ⅰ은 교목층에서 소나무가 우점하고, 관목층과 초본층에서는 주걱댕강나무가 우점하는 가운데 털진달래, 철쭉, 쇠물푸레나무의 경쟁 관계가 지속될 것으로 판단된다.
군집 Ⅱ (신갈나무-주걱댕강나무 군집)는 교목층에서 신갈나무(I.P.: 73.2%), 굴참나무(I.P.: 19.9%)가 주요 출현종이었으며(Table 2), 굴참나무의 경우 고사(DBH 8 ~ 14 cm, 11개체)되거나 가지가 말라 들어가는 개체(DBH 8 ~ 20 cm, 9개체)가 나타나는 등 세력이 약화 징후를 보여, 천이계열 진행에 따라 신갈나무가 더욱 우점 할 것으로 판단된다. 아교목층에서도 신갈나무(I.P.: 35.6%)가 우점하는 가운데, 주걱댕강나무(I.P.: 17.7%), 쇠물푸레나무(I.P.: 10.3%), 털진달래(I.P.: 7.3%)가 출현하였다. 관목층에서는 주걱댕강나무(I.P.: 63.5%)가 우점을 보였고, 털진달래(I.P.: 18%), 쇠물푸레나무(I.P.: 7.5%)와 생육 경쟁 관계가 이어질 것으로 판단된다(Table 2). 초본층은 전반적으로 주걱댕강나무(I.P.: 36.3%)가 우점을 보이는 가운데, 조릿대가 다수 출현하고 있는 조사구(12, 17, 20)의 경우 조릿대 확산과 주걱댕강나무 생육압박 정도에 대한 지속적인 관찰이 요구된다.
군집 Ⅲ (졸참나무-주걱댕강나무 군집)은 교목층에서 졸참나무(I.P.: 43%)와 굴참나무(I.P. : 36.9%)가 주요 출현종이었고, 아교목층에서는 사람주나무(I.P.: 25.7%)가 우점하는 가운데 철쭉(I.P.: 10%), 개옻나무(I.P.: 7.8%), 대팻집나무(I.P.: 7.3%)가 함께 출현하였다(Table 2). 관목층은 주걱댕강나무(I.P.: 55.9%)가 우점종이었으며, 철쭉(I.P.: 8.8%), 사람주나무(I.P.: 6.9%), 털진달래(I.P.: 6.4%) 등이 함께 출현하여 생육 경쟁 관계를 형성하고 있었다. 초본층에서도 주걱댕강나무(I.P.: 58.8%)가 우점종이었으며, 조릿대(I.P.: 11.4%)가 부분적으로 출현하였다(Table 2). 군집 Ⅲ은 교목층에서 졸참나무와 굴참나무가 우점하고, 아교목층 이하 모든 층위에 출현하는 사람주나무, 철쭉, 털진달래는 장래 관목층 이하 우점종인 주걱댕강나무의 생장에 직접적인 영향을 초래하는 요인이 될 것으로 판단된다.
군집 Ⅳ (개서어나무 아군집)는 교목층에서 개서어나무(I.P.: 73.5%)가 높은 우점을 보이는 가운데 신갈나무(I.P.: 26.5%)가 함께 층을 이루었다(Table 2). 아교목층에서는 철쭉(I.P.: 32.2%), 개서어나무(I.P.: 19.5%), 사람주나무(I.P.: 18%), 쇠물푸레나무(I.P.: 16%) 등이 출현하였고, 관목층에서는 주걱댕강나무(I.P.: 70.1%)가 높은 우점을 보였으며, 쇠물푸레나무(I.P.: 6.3%), 사람주나무(I.P.: 4.6%), 생강나무(I.P.: 4.4%) 등이 더불어 출현하였다. 초본층에서도 주걱댕강나무(I.P.: 45.2%)가 우점하는 가운데, 쇠물푸레나무(I.P.: 12.7%), 생강나무(I.P.: 11.4%), 철쭉(I.P.: 8.1%) 등이 출현하였다(Table 2). 군집 Ⅳ는 산복부의 계곡형태 미세지형으로 다른 군집과는 분류군에서 차이가 많아 가장 먼저 분리 되었으며, 교목층에서는 개서어나무의 우점이 더욱 확대될 것으로 판단되며, 관목층 이하에서는 주걱댕강나무의 우점이 당분간은 유지될 것이나, 낮은 광 환경에서도 생육이 양호한 쇠물푸레나무, 생강나무, 철쭉 등의 생장에 따라 경쟁관계를 형성해 나갈 것으로 예상된다.
군집별 평균상대우점치(M.I.P.)는 군집 Ⅰ에서 소나무가 35.9%로 가장 높았고, 관목층과 초본층의 우점종인 주걱댕강나무는 15.6%이었으며, 아교목층과 관목층의 분포 비중이 높은 쇠물푸레나무 9.8%, 털진달래 6.9%로 분석되었다(Table 2). 군집 Ⅱ는 주로 능선부나 상대적으로 해발고도가 높은 사면 상부에 위치(평균 해발고도 472.71 m)하는 집단으로 신갈나무의 평균상대우점치(M.I.P.)가 33.6%로 가장 높았으며, 관목층과 초본층 우점종인 주걱댕강나무는 26.1%, 아교목층과 관목층의 분포 비중이 높은 털진달래 7.3%, 쇠물푸레나무 6.1% 순이었다(Table 2). 군집 Ⅲ은 상대적으로 해발고도가 낮은 사면하부에 위치(평균 해발고도: 239.5 m) 하는 집단으로 졸참나무 19.2%, 주걱댕강나무 18.2%, 굴참나무 16.1% 순으로 나타나 특정 분류군이나 특정 층위에 치우치지 않은 집단으로 구분되었다(Table 2). 군집 Ⅳ는 사면상부의 계곡형 포행토가 진행 중인 곳으로 교목층의 개서어나무가 35.2%로 가장 높았으며, 관목층과 초본층의 우점종인 주걱댕강나무는 18.5%로 나타났다(Table 2).
이상 군집별 분석 결과를 종합하면, 천성산 주걱댕강나무 개체군 내 상층식생은 소나무, 참나무류, 개서어나무가 우점을 하고 있으나 식생 천이계열에 따라 신갈나무, 개서어나무 군집으로 천이가 진행될 것으로 판단되며, 주걱댕강나무는 다층구조의 층위에서 관목층과 초본층에 우점종으로 분포하는 관목으로 당분간은 현재와 같은 하층식생 범위 우점을 유지하겠으나, 장래 같은 층위에 분포하고 있는 음지 적응성이 높은 쇠물푸레나무, 털진달래, 철쭉, 사람주나무, 조릿대 등의 세력 확산 시에는 생육 경쟁관계가 촉진될 것으로 보여 지속적인 모니터링과 보호관리가 요구된다.
Table 2.
Importance percentage of the major species in the Diabelia spathulata population
유사도 분석
조사구의 지형에 따른 일조시간, 온도, 토양의 함수량, 전질소 함량, 가용성 인의 함량 등 환경요인은 종구성과 유사도에 많은 영향을 준다(Cho and Yoon, 1996). 주걱댕강나무 개체군에 대한 군집별 유사도(Sorensen, 1948)를 분석한 결과(Table 3), 군집 Ⅰ과 군집 Ⅱ는 69.14%, 군집 Ⅱ와 군집 Ⅲ은 59.41%, 군집 Ⅰ과 군집 Ⅲ은 54.17%로 비교적 이질성이 크지 않은 것으로 나타났으며, 군집 Ⅲ과 군집 Ⅳ는 35.14%, 군집 Ⅱ와 군집 Ⅳ는 33.9%, 군집 Ⅰ과 군집 Ⅳ는 33.33%로 다소 이질적인 것으로 분석되었다(Table 3). 이들 중 군집 Ⅰ (소나무 군집)과 군집 Ⅱ (신갈나무-주걱댕강나무 군집), 군집 Ⅲ (졸참나무-주걱댕강나무 군집)은 출현 종수 등에서 차이가 크지 않았으나, 군집 Ⅳ (개서어나무 군집)의 경우 다른 군집에서 출현하지 않는 회잎나무, 둥근잎천남성, 비늘고사리 등 종구성과 생육 개체수의 차이로 인해 군집 Ⅳ와 결부되어 유사도가 모두 낮게 나타난 것으로 판단된다.
이를 통해 군집 사이의 유사성 정도는 개체군 구조에 크게 의존되고 있음을 확인할 수 있었다(Jokimäki and Kaisanlahti-Jokimäki, 2003). 또한 전체 군집 유사도지수 범위가 33.33 ~ 69.14로 나타나 주걱댕강나무 개체군은 식생 천이가 진행 중인 것으로 분석되었다.
Table 3.
Similarity index (%) between communities
| Community | Ⅰ | Ⅱ | Ⅲ |
| Ⅱ | 69.14 | ||
| Ⅲ | 54.17 | 59.41 | |
| Ⅳ | 33.33 | 33.9 | 35.14 |
종다양도
주걱댕강나무 개체군의 종다양도(Shannon-wiener)를 군집별로 분석한 결과(Table 4), 4개 군집의 종다양성지수(Species diversity index)는 0.6615 ~ 0.8267 범위로 나타났다. 종다양도(H')와 최대종다양도(H'max)로 산출되는 균재도(J’)는 그 값이 1에 가까울수록 종별 개체수가 편중되지 않고 균일한 상태를 나타내는 것으로(Brower and Zar, 1977), 주걱댕강나무 개체군의 군집별 출현 식물 종수는 16 ~ 58종 범위(군집 Ⅰ-38종, 군집 Ⅱ-43종, 군집 Ⅲ-58종, 군집 Ⅳ-16종)였고, 균재도는 0.5260 ~ 0.6866으로 식생의 균일성이 그다지 높지 않은 것으로 분석되었다(Table 4).
이는 주걱댕강나무 생립지의 교목층을 우점하는 특정 분류군(군집 Ⅰ-소나무 78.1%, 군집 Ⅱ-참나무류 93.1%, 군집 Ⅲ-참나무류 93.2%, 군집 Ⅳ-개서어나무 73.5%)의 편중이 종간 균등성을 저해한 것으로 보인다. 우점도(D)의 경우 0.9 이상일 때에는 특정 종이 압도적인 우점을 보이며, 0.3∼0.7 사이이면 1종 또는 2종이 우점하며, 0.1∼0.3일 때에는 여러 종이 우세를 나타내는데(Whittaker, 1956), 본 조사에서 우점도가 0.3134 ~ 0.4740 범위로 나타나 특정 종 우점에 근접하는 식생형을 보였다.
Table 4.
Species diversity indices of community
| Community | No. of species | H' | H'max | J' (evenness) | D (dominance) |
| Ⅰ | 38 | 0.7938 | 1.2187 | 0.6526 | 0.3474 |
| Ⅱ | 43 | 0.6615 | 1.2528 | 0.5260 | 0.4740 |
| Ⅲ | 58 | 0.7638 | 1.3417 | 0.5657 | 0.4343 |
| Ⅳ | 16 | 0.8267 | 1.2041 | 0.6866 | 0.3134 |
생육 환경
토양
토양온도는 평균 16.71 ℃로 고도에 반비례하였고, 지형과 방위에 따라 차이를 보였다(Table 1). 토양습도는 평균 16.64%로 능선부, 사면 중・상부, 볕이 잘 드는 곳에 주로 분포하는 입지특성과 함께 토질과 경사・방위에 따라 다양한 측정값을 보였다. 이는 관목성이면서 주걱댕강나무와 비슷한 개체군을 형성하는 꼬리말발도리 자생지 중 식물구계학적으로 남부아구에 속하는 밀양지역에서 측정된 꼬리말발도리 집단의 평균 33.4% (Jung et al., 2016)와 차이를 보였으며, 소백산 삼가지구 등산로 구간 평균 26.37% (Park et al., 2020), 지리산 세석지구 북사면 평균 21.7% (Noh et al., 2018)에 비하면 다소 건조한 지역으로 판단된다.
주걱댕강나무 생육지에 대한 토양의 물리・화학적 특성을 분석한 결과(Table 5), 심토는 갈색 내지 암갈색의 미사질양토였으며(National Institute of Agricultural Sciences, 2020), 토양산도는 pH 4.2 ~ 4.9 범위로 모든 조사구에서 강산성을 띠었다. 이는 우리나라 산림토양 평균산도(pH 4.88)와 유사한데(Kim et al., 2018), 낮은 산도는 식물이 이용할 수 있는 인산, 칼슘, 마그네슘 등을 불용화 시켜 유효도 또한 낮은 결과로 나타났다. 특히 식물 지하부발달에 영향을 미치는 유효인산의 경우 수목생육 적정범위(100 mg kg-1 이상)보다 많이 부족한 평균 19.3 mg kg-1로 분석되었다. 주걱댕강나무 개체군 전체 조사구에서 추출한 유기물 함량(%)은 7.7 ~ 19.6 범위로, 우리나라 산림토양의 유기물함량 평균 4.1보다 높은 값을 보였다(Kim et al., 2018). 토양특성에 중요한 영향을 미치는 유기물(Organic matter)은 많은 음전하를 가지기 때문에 토양의 양이온 교환 능력을 증대시키고, 토양의 통기성과 투수성・보수력을 좋게 하므로 유기물 함량이 높으면 건강한 식생 형성에 도움이 된다(Brady, 1990). 또한 식물의 생리작용을 촉진하는 물질을 공급하는 원소로서 토양유기물함량과 상관관계가 높은 전질소 함량(%)은 0.44로 나타나 우리나라 산림토양의 평균 전질소 함량 0.17보다 높았으며, 수목생육 적정범위(0.25 이상)를 상회하는 값을 보였다.
이상에서 나타난 주걱댕강나무 개체군의 토양특성은 유기물과 전질소 함량은 높게 나타난 반면, 매우 낮은 토양산도는 식물생육에 필요한 유효 영양소의 불균형을 가져올 수 있어 토성변화에 대한 지속적인 관찰이 필요하다.
Table 5.
Soil characteristic in each studied plots
빛 환경
수목의 생리적 특성과 밀접한 관련성이 있는 수관열림도(Canopy Openness)는 조사구 3에서 27.65%로 가장 높았으며, 조사구 12에서 26.2%, 조사구 7에서 22.15% 순으로 나타났고, 조사구 17과 19에서는 각각 7.59%, 9.86%로 상대적으로 낮았으며, 평균 수관열림도는 18.56 ± 4.79%였다(Table 6).
Table 6.
Light characteristic in each studied plots
| Plot No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | Mean |
|
Canopy Openness | 17.7 | 11.8 | 32.4 | 28.3 | 20.6 | 22.0 | 22.5 | 21.4 | 21.9 | 30.0 | 31.5 | 30.2 | 20.7 | 19.2 | 15.5 | 24.0 | 8.5 | 23.0 | 8.3 | 27.6 | 21.86 |
이는 식물구계학적으로 남부아구에 속하는 울산지역에서 측정된 소관목 특성을 갖는 거문도닥나무 집단의 평균 45.23 ~ 52.87%보다는 낮았고(Yoon et al., 2014), 관목형 수형을 갖는 꼬리말발도리 집단의 평균 15.5%와 유사하였다(Jung et al., 2016).
엽록소 함량
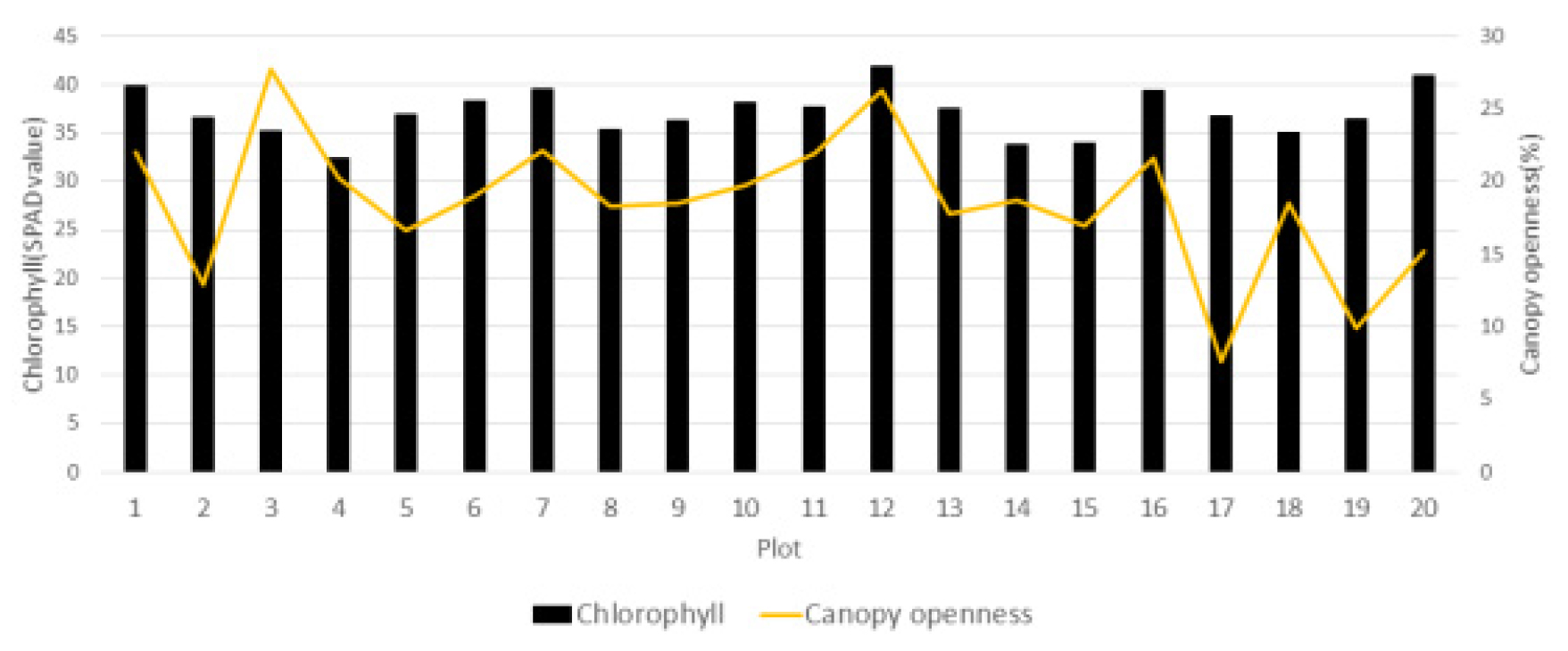
주걱댕강나무의 엽록소 함량(SPAD value)은, 조사구 12에서 41.9로 가장 높은 값을 보였고, 조사구 4와 17에서는 각각 32.4, 33.1로 상대적으로 낮은 값을 나타내었으며, 20개 조사구의 평균값은 36.74 ± 2.80으로 산출되었다(Fig. 4).
이는 높이 1 ~ 4 m까지 자라는 낙엽활엽관목으로 전라도, 경상도 지역에 자생하는 정금나무 10개 집단의 측정값(35.0 ± 5.9)과 유사하며(Kim et al., 2012), 국립수목원(3개소)과 경상북도 팔공산(1개소) 꼬리말발도리 자생지의 엽록소 지수(25 ~ 40)와도 유사한 경향을 보였다(Jung, et al., 2020). 수관열림도와 상관에서는 대체로 수관열림도가 높아질수록 엽록소 지수도 높아지는 경향을 보였는데(Fig. 4), 이는 내음성이 강한 수종은 빛 환경에 대한 적응반응으로 수관열림도가 낮아질수록 수광량과 빛의 이용효율을 높여 엽록소 함량이 증가하는 반면, 상대적으로 광포화점이 높은 양수는 수관열림도가 높아질수록 엽록소 함량이 증가하는 일반적인 방향성에 부합하는 결과를 보였다(Hansen et al., 2002; Lee, 1993).
개체군 특성
주걱댕강나무 개체군 20개 조사구(개소당 면적: 100 ㎡)에서 확인된 주걱댕강나무의 총 개체수는 3,270개체였으며, 조사구 12에서 486개체로 가장 많았고, 조사구 7은 344개체, 조사구 3은 276개체 순으로 나타났으며, 조사구 2와 14에서는 각각 51개체로 가장 적었다(Table 7). 개체밀도(개체수/ ㎡)는 조사구 12에서 4.86으로 가장 높았고, 조사구 2와 14에서 각각 0.51로 가장 적은 값을 보였다.
Table 7.
Density and flowering rate of Diabelia spathulata by studied plots
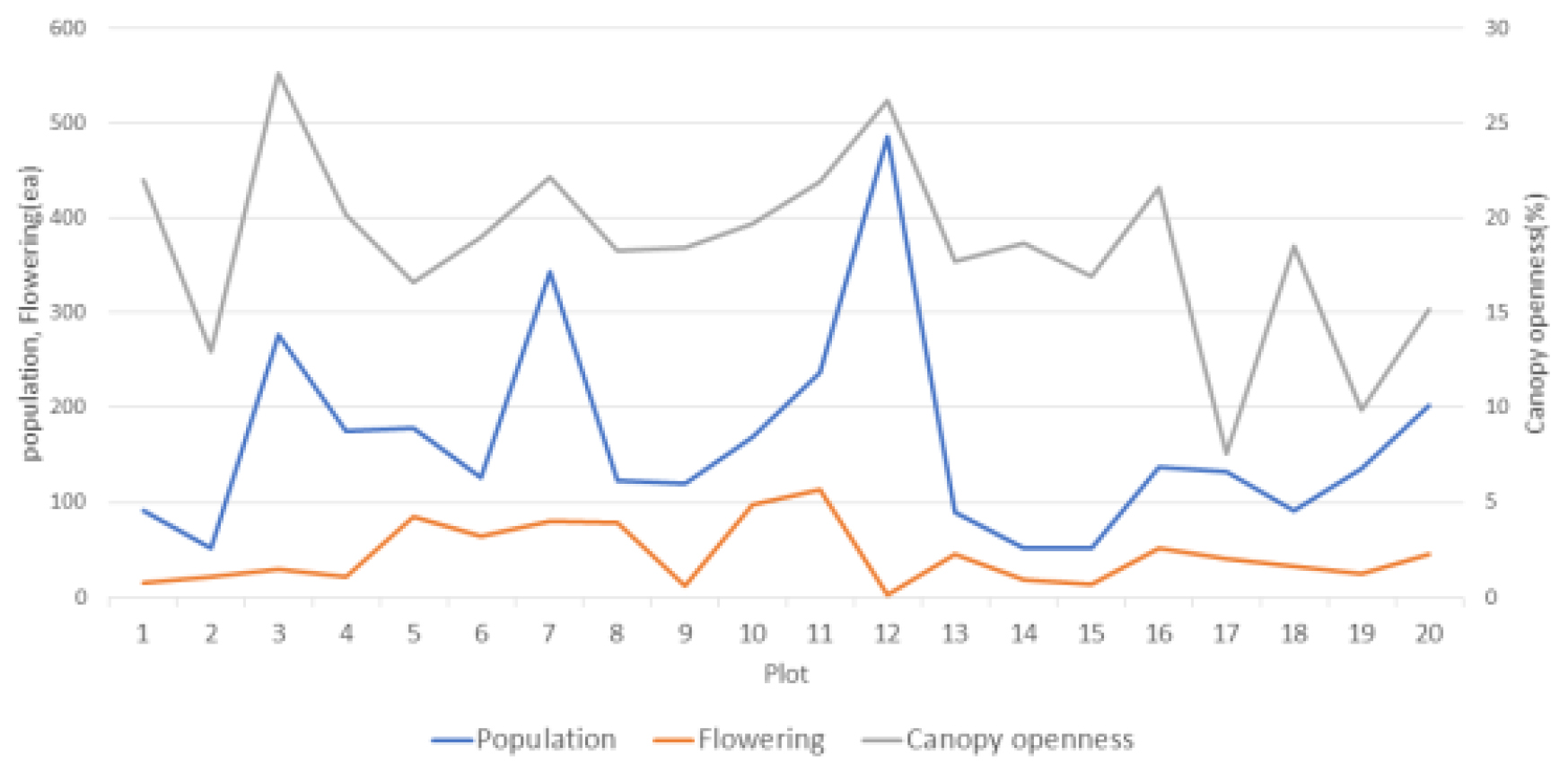
주걱댕강나무 분포는 초본층에서 아교목층 일부까지 최고수고 3.4 m까지 형성되어 있었으나, 평균 수고가 1.1 m로 주로 관목층 이하 높이에 분포하고 있는 것으로 나타났다. 20개 조사구의 개체수 대비 평균 개화율은 27.37%였으며, 조사구 8에서 64.23%로 가장 높았고, 조사구 13은 51.69%, 조사구 6은 50.79% 순으로 나타났으며, 조릿대가 초본층을 우점하는 조사구 12는 0.41%로 가장 낮게 나타났다(Table 7). 개화 개체의 수고는 최소 0.3 m에서 최고 3.4 m 범위였으나 개화가 집중되는 수고 분포 범위는 1.0 ~ 1.8 m (평균수고 1.39 m) 사이였다. 이는 주걱댕강나무가 주로 관목층(H: 0.8 ~ 2 m)까지 분포하되 아교목층(H: 2 ~ 8 m)에서는 사람주나무, 쇠물푸레나무, 털진달래, 철쭉 등 음지에서 세력 확장이 강한 수종들에 의한 두터운 피음이 개화를 억제하는 요인으로 작용했기 때문인 것으로 판단된다. 이상의 결과를 종합해보면, 주걱댕강나무는 교목층이나 아교목층이 없어 수관열림도는 높고 토심이 깊은 곳에서 많은 개체수를 확인 할 수 있었다. 이는 개체의 특성상 수고가 제한적으로 자라기 때문인 것으로 판단되며, 개화 정도는 개체가 일정수준 이상(H: 1.0 m)으로 생장하고 분포층위 내 피음이 낮을수록 개화율이 높은 것으로 분석되었다(Fig. 5).
본 조사에서 확인 한 주걱댕강나무 집단의 개체 확장 형태는 주로 표토와 가까운 3 ㎝ 범위 내에서 뿌리줄기를 횡방향으로 뻗으면서 지상줄기를 내어 군락을 이루는 영양번식 형태를 취하고 있었는데, 이는 매우 낮은 결실율(Kim and Kim, 2011)에 따라 종자번식방법으로는 원활한 개체군 재생산이 어렵기 때문에 채택된 생존양식으로 판단된다. 따라서 향후 주걱댕강나무 개체군 확대 또는 대체 서식지 조성을 위해서는 영양번식 방법 중 근삽이나 분주 그리고 녹지삽목이 잘 되는 특성(Noboru, 1979)을 고려하여 다양한 증식방법에 대한 추가적인 연구로 최적의 대량증식을 위한 방안도 모색해 나가야 할 것으로 판단된다.
상관분석
주걱댕강나무 개체군의 환경요인과 생육특성 간 상관관계를 분석한 결과(Table 8), 해발고도는 경사와 종다양도・균재도와 음의 상관을, 우점도와는 양의 상관 관계를 보였다. 이는 주걱댕강나무 생육지의 해발고도가 높을수록 경사가 완만한 능선이나 사면상부 분포가 많았고, 고도에 따라 참나무류와 개서어나무 등 특정 분류군의 높은 우점에 기인한 것으로 판단된다. 또한 경사는 교목층 피도와 양의 상관을, 초본층 피도와는 음의 상관으로 나타났다. 이는 경사가 급할수록 수광 범위가 좁아져 상층부 수관은 광량과 세력이 유지되는 반면, 상대적으로 광량이 부족해지는 지표층의 발달은 미흡하기 때문인 것으로 판단된다. 수관열림도는 엽록소함량과 양의 상관을 보여 양지환경을 선호하는 주걱댕강나무 특성(Ishii et al., 2010)과 부합하였다. 한편 식물의 생장에 큰 영향을 미치는 유기물은 전질소와 강한 양의 상관관계를 형성하였고(Park, 2019), 종다양도는 균재도와 양의 상관을, 우점도와는 음의 상관을 보였으며, 이는 일반적인 식물 군집 내 종수와 종별 개체수에 따른 상관관계 경향과 유사하게 나타났다.
Table 8.
Correlation analysis of environmental factors and vegetation characteristics in Diabelia spathulata habitats
zsignificance at 5% level, ysignificance at 1% level, xAT: Altitude, wSD: Slop degree, vCO: Canopy openness, uCC: Chlorophyll content, tNS: No. of species, sCT: Coverage of tree, rCU: Coverage of understory, qCS: Coverage of shrub, pCH: Coverage of herb, oOM: Organic matter, nTN: Total nitrogen, mDIV: Species diversity, lEVE: Evenness, kDOM: Dominance.
적 요
주걱댕강나무는 세계적으로 중국 남부・일본 혼슈 이남에 분포하며, 국내에서는 유일하게 경상남도 양산시 천성산에만 분포하는 분류군으로, 2003년도에 국내 야생 존재가 보고된 이래 지금까지 식생구조나 생육특성 등 일체 선행연구는 찾아볼 수가 없었다. 이에 본 연구는 주걱댕강나무 집단의 분포범위, 생육환경과 개체 특성에 대한 분석을 통해 주걱댕강나무의 적절한 현지 내・외 보전관리를 위한 기초자료를 확보하기 위해 수행하였다. 이번 조사를 통해 확인한 주걱댕강나무 개체군의 범위는 지리적으로 북위 35°24' 58"에서 35°26' 35", 동경 129°05' 43"에서 129°07' 04" 사이에 분포하고 있었으며 해발고도는 98 ~ 592 m, 반경 1.8 ㎞ 범위 내 능선과 사면, 계곡부 등 사방에 걸쳐 햇빛이 어느 정도 확보되는 곳에서 소규모 군락형태로 관찰되었다. 주걱댕강나무 개체군의 식생구조는 소나무군집, 신갈나무-주걱댕강나무군집, 졸참나무-주걱댕강나무군집, 개서어나무아군집으로 분류되었고, 교목층을 우점하는 특정 분류군(소나무, 참나무류, 개서어나무)의 편중으로 종간 균등성은 낮았으며, 우점도 또한 특정 종 우점에 근접하는 식생형을 보였다. 주걱댕강나무 개체군 20개 조사구에서 측정한 전 분류군의 평균 수고는 교목층 8 ~ 13 m, 아교목층 3.4 ~ 6 m, 관목층 1.2 ~ 1.7 m, 초본층 0.2 ~ 0.7 m 범위의 다층구조로 형성되어 있었다. 토양 유기물 함량은 비교적 높은 특징을 보였으나 모든 조사구에서 토양산도(pH) 4.2 ~ 4.9의 강산성을 띠고 있어 식물생육에 요구되는 유효인산, 마그네슘 등 필수 원소는 불균형으로 나타났다. 또한 조사지 내 주걱댕강나무의 엽록소 함량은 36.74 ± 2.8로 식물구계학적으로 남부아구에 속하고, 주로 하층식생으로 분포하는 정금나무 집단의 35 ± 5.9와 유사하였으며, 수관열림도가 높아질수록 비례하여 엽록소 함량도 증가하는 양의 상관을 나타내었다. 주걱댕강나무 개체군은 입지특성과 층위에 따라 신갈나무, 철쭉, 털진달래, 쇠물푸레나무, 생강나무와 높은 양의 식생상관을 보였으며, 다층구조 수관층위에서 최고수고 3.4 m까지 형성되어 있었으나 평균 수고가 1.1 m로 주로 관목층과 초본층에 우점종으로 분포하였다. 주걱댕강나무의 개화 개체 수고는 최소 0.3 m에서 최고 3.4 m 범위였으며, 개화가 집중되는 수고는 1.0 ~ 1.8 m (평균수고 1.39 m) 사이였고, 평균개화율은 27.37%였다. 현지 내 보전에 있어서 임지 내 수목의 생장은 기후, 미세지형, 토양조건 등에 의한 입지환경의 영향을 크게 받는데, 이러한 입지환경 중 수목의 생육에 결정적인 역할을 하는 요소는 광도와 온・습도 등 기후적인 인자이다. 조사 대상지에서 주걱댕강나무 치수가 많은 곳은 교목층이 없거나 아교목층이 없어 상대적으로 수광이 용이한 곳이었으며, 양지환경을 선호하는 주걱댕강나무 특성을 감안할 때 교목층을 우점하고 있는 참나무류와 소나무, 아교목층과 관목층의 사람주나무, 쇠물푸레나무, 털진달래, 철쭉 그리고 초본층의 조릿대 등과 생육 경쟁관계 형성 또는 이들로 부터 피음을 받을 것으로 보여 주걱댕강나무 집단의 개체수 감소를 방지하고 주걱댕강나무가 선호하는 광 환경을 유지하기 위해서는 층위별로 밀도조절 등 관리가 요구되며, 이를 위해 지속적인 모니터링과 주변식물 종들의 적절한 제어가 필요할 것으로 판단된다.
