

서 언
보전생물학(conservation biology)은 생태계 내 생물종과 환경간의 상호관계를 파악하고 이를 보존하는 것이 목적이다. 즉, 지구상의 생물다양성 목록을 구축하고, 인간과 생태계의 상호관계 연구와 생물종 절멸(extinct)을 방지 및 건강한 생태계 기능을 증진하는 것이 보전생물학의 핵심목적이다(Wilson, 1992). 생태계 구성요소는 지구환경변화에 의한 영향을 받고 있다. 또한, 난·온대성 식물의 분포는 확산되나 한대성 식물의 분포는 축소되고, 저지대 식물이 고지대로 이동하고 있다(Kullmann. 2002; Meshinev et al., 2000). 이에 지구환경변화에 따른 생물종분포 연구가 중요시 되고 있으며 멸종위기 식물을 대상으로 종보전의 필요성이 대두되고 있다(Golicher et al., 2008). 특히 고산 지대에 생육하는 침엽수종의 생육지 감소로 정밀한 모니터링이 필요하다(Kong, 2002). 국외 Abies 속의 아고산 침엽수종 쇠퇴원인에 대한 연구는 풍해, 수분스트레스 등에 의한 것이라는 의견이 지배적이다(Germino et al., 2002; Hasegawa and Mori, 2007; Lee and Kim, 2007; Mori et al., 2004).
전나무속(Abies)은 세계적으로 52종이 분포하고 있으며, 한국에서는 전나무(Abies holophylla Maxim.),구상나무(Abies koreana E.H. Wilson), 분비나무(Abies nephrolepis (Trautv. ex Maxim.) Maxim.)로 3종이 생육하고 있다. 구상나무는 한국에 자생하는 특산식물로 주로 또한 기후변화 취약함이 인정되어 보전가치가 있는 식물종 중 하나이다(Kong, 2002). 특히 한라산 일대에는 대규모의 구상나무 군집이 형성되어있어 구상나무림의 동태를 살피는 대표적인 연구대상지로 가장 많은 연구가 진행되어 있었으며. 특히 국내 구상나무의 생육지 감소에 따른 쇠퇴원인 규명에 대한 연구가 활발히 이루어 졌는데, 수분스트레스와 풍해에 의한 물리적 피해 등 여러 가지 가설이 제시되어 왔다(Kim and Choo, 2000; Kim et al., 2007; Kong, 1998, 2006; Lim et al., 2006; Song et al., 2010). 그러나 40여년 간의 구상나무의 연구가 진행되어 왔으나, 명확한 쇠퇴와 고사에 대한 원인은 분명하게 규명되지 않았으며, 구상나무 개체군동태에 대한 임분구조 등의 연구가 한 산지 또는 특정지역에 국한되어 있어 전반적인 구상나무의 생육환경과 종조성에 대한 상호관계에 대한 데이터가 미흡한 실정이다(Koo and Kim, 2020). 2000년도 구상나무를 대상으로 덕유산, 지리산, 한라산을 대상으로 지역별 개체군 구조의 차이와 환경요인간의 상관관계에 대하여 연구한 바가 있으나 그 이후 여러 자생지를 대상으로 비교한 연구는 미흡하다(Kim and Choo, 2000). 본 연구의 목적은 국내 대표적인 구상나무 자생지인 덕유산, 지리산, 한라산을 대상으로, 종조성과 토양이화학적성질과 경관지수를 이용하여 구상나무 분포와 환경요인간의 종합적인 상호관계를 다각적으로 분석하여 향후 구상나무 보전을 위한 기초자료를 마련하는 것이다.
재료 및 방법
조사지현황
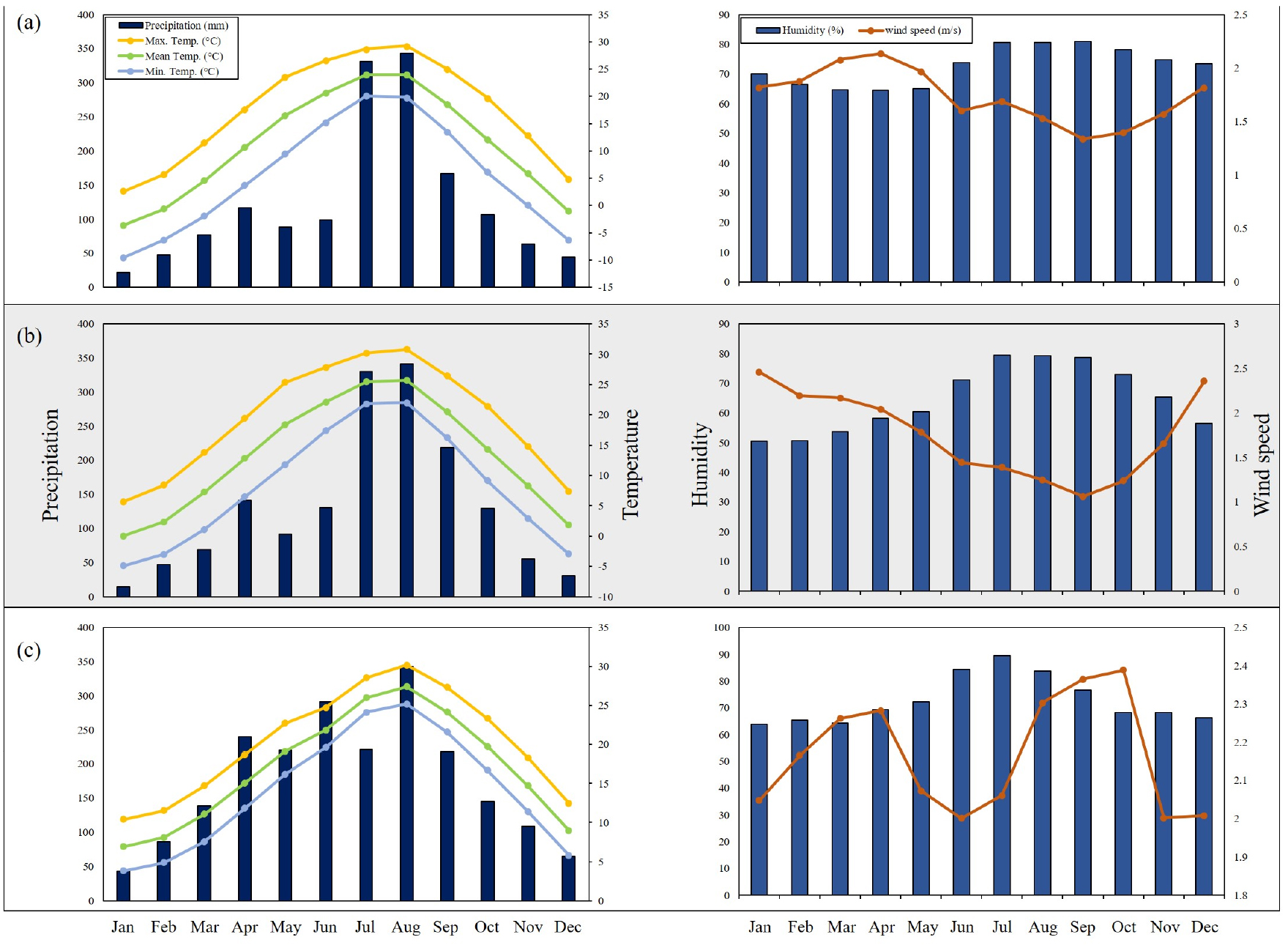
본 연구대상지는 덕유산(1,614 m), 지리산(1,915 m), 한라산(1,947 m)에 분포하고 있는 구상나무림을 대상으로 실시하였다. 해발고도 1,000 m 이상의 지점 대상으로 표본점 조사를 시행하였으며, 덕유산 24개소, 지리산 61개소, 한라산 36개소로 총 121개소의 표본점 조사를 시행하였다(Fig. 1). 조사지 좌표는 동경 126° 30′30.3″~ 127° 45′18.46″, 북위 33°20′54.1″~ 35° 51′ 56.4″에 위치하고 있으며 행정구역상 위치는 덕유산의 경우 전라북도와 경상남도, 지리산은 전라남·북도, 경상남도 그리고 한라산은 제주특별자치도에 위치하고 있다. 기후환경은 Korea Meteorological Administration에서 제공하는 최근 10년간(2011 ~ 2020) 기상자료를 이용하였다(Korea Meteorological Administration, 2021). 연구대상지와 가장 근접한 기상관측소의 기상데이터를 이용하여 덕유산은 장수기상관측소, 지리산은 산청기상관측소, 한라산은 서귀포기상관측소로 선정하였다. 최고기온, 최저기온, 평균기온, 강수량과 평균풍속 및 상대습도를 분석하였다. 월최고기온의 평균은 덕유산 17.3℃, 지리산 19.2℃, 한라산 20.2℃으로 나타났으며, 월평균기온의 평균은 덕유산 10.9℃, 지리산 13.2℃, 한라산 16.9℃로 나타났으며, 월최저기온의 평균은 덕유산 5.3℃, 지리산 8.1℃, 한라산 14.0℃로 나타났다. 연 평균강수량은 덕유산 1,506.0 ㎜, 지리산 1,597.8 ㎜, 한라산 2,119.7 ㎜로 한라산에서 가장 많은 강수량으로 조사되었다. 평균풍속의 경우 덕유산 1.73 m/s, 지리산 2.35 m/s, 한라산 2.16 m/s로 나타났으며, 상대습도는 덕유산 7.7%, 지리산 56.5%, 한라산 72.6%로 지리산에서 다소 건조한 상대습도로 나타났다(Fig. 2).
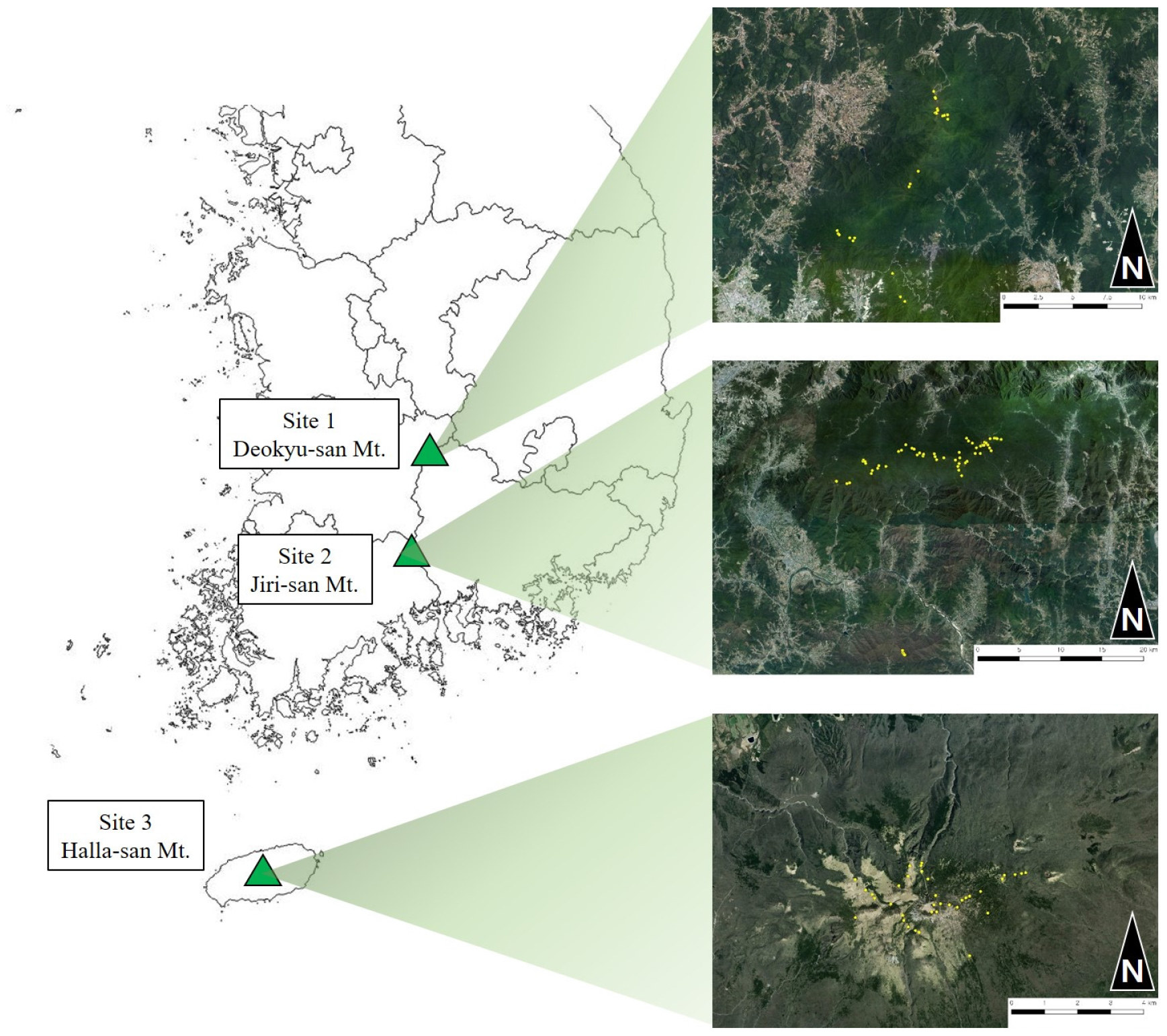
조사구설치 및 조사방법
표본점은 원형방형구를 설치하였으며, 반지름 11.3 m로 400 ㎡ 내 매목조사 및 식생조사를 시행하였다. 이는 산림식생형에서 수관을 이루는 교목성 수종군락에서의 최소 조사구면적인 200 ~ 500 ㎡의 기준(Kim and Lee, 2006)을 고려하였다. 조사기간은 2019년 6월부터 2020년 10월까지 조사하였다. 매목조사는 흉고직경 6 ㎝ 이상의 모든 임목에 대하여 흉고직경, 수종명을 조사하였다. 식생조사는 수직적 위치를 고려하여 층위별로 목본층(교목․아교목․관목층)과 하층(초본층)으로 구분하여 목본층의 경우 2 ㎝ 이상 개체목의 흉고직경(DBH)을 측정하였고, 초본층의 피도는 Braun-Blanquet (1964)의 통합우점도를 이용하였다.
분석방법
토양 이화학적 성질분석을 위하여 표준지 조사가 이루어진 121개소 가운데 77개소의 표준지에 대하여 토양시료를 채취하였다. 채취장소는 암석노출도가 높지 않고 토양이 발달된 지역에 한하여 채취하였으며, 유기물층을 제거한 후 0 ~ 20 ㎝의 깊이에서 시료를 채취하였다.
채취한 토양은 밀봉한 상태로 분석실로 운반하여 실내에서 약 2주간 풍건 후 2 ㎜와 0.02 ㎜의 체에 통과시켜 분석용 시료를 제작하였다. 토양의 물리적 성질인 토성은 비중계법으로 측정하였다. 토양pH 및 전기전도도는 풍건세토 10 g과 증류수 50 ㎖를 1:5로 만든 후 pH-meter를 사용하여 측정하였다. 양이온치환용량(CEC)은 Brown’s Method법으로 분석하였으며, 치환성 양이온의 경우 불꽃반응 법으로 분석하였다.
경관지수 도출을 위하여 Arcgis 10.1 ver.를 이용하였으며, 지형습윤지수(TWI, Topographic Wetness Index), 지형지수(TPI, Topographic Position Index) 그리고 Solar radiation으로 3가지의 경관지수를 추출하였다. 본 경관지수는 수치표고모델(DEM, Digital Elevation Model)을 기반으로 추출하였다. 획득한 DEM 원자료는 DEM 생산 과정에서 오류가 발생할 수 있어 오류 격자 추출 후, 주변 격자를 이용하여 보정을 수행하는 Fill 툴을 통해 오류가 제거된 DEM을 생산하였다. 지형습윤지수는 대면적 범위의 토양 수분 분포 패턴의 정보를 제공하는 지도화에 소요되는 높은 수준의 비용을 저감하기 위해 원격탐사기법을 활용한 지도화 방법으로서, 지표면 지형이 지하수의 깊이와 유로에 영향을 미치는 주요한 요인이라는 가정에 기초한다(peterson et al., 2009). 이러한 가정과 함께 TWI는 특정 지점의 국부 경사면 상에서 특정 지점을 향해 흐르는 수분의 측정치로 정의된다. 이에 국부 경사 수준이 증가하거나 집수 면적이 낮을수록 토양 수분이 적으며, 반대로 국부 경사 수준이 낮아지거나 집수면적이 높을수록 토양이 머금은 수분 조건이 양호함을 의미한다(Petroselli et al., 2013). TWI를 구하는 공식은 ln (A/tanB) 로 A는 집수면적(contributing area)이고 B는 경사도이다. DEM을 바탕으로 격자들 간의 상대적인 해발고도 고저에 따라 유수 방향을 결정하기 위해 flow direction 툴을 이용하여 유로 방향 지도를 생산하였다. 그리고 보정된 DEM을 이용하여 TWI의 주요 요소인 국부 경사지도를 생산하였다. 다음 단계에서는 유로 방향 지도를 바탕으로 각 격자별로 유수 축적 수준을 계산하기 위해 Flow accumulation 툴을 통해 격자별 유수 축적 지도를 생산하였다. 마지막 단계로 국부 경사지도와 격자별 유수 축적 지도를 바탕으로 Raster calculator 툴을 통해 TWI 계산식을 이용하여 TWI 지도를 제작하였다. TPI는 지형적 특성을 이용하여 능선부 및 계곡부 형태의 정량적 수치를 제공하며, 지수값이 음수일수록 계곡부(오목한 지형)이며, 양수일수록 능선부(볼록한 지형)의 형태를 나타내었다.일사량은 미터당 와트시간 단위미터당 와트시간 단위(WH/㎡)를 가지며 높을수록 많은 일사량을 받는 지형이며 반구형 가시권 알고리즘 기반으로 도출하였다. 이는 ArcGIS 내 Arctoolbox의 ‘Area Solar Radiation Tool’ 사용하여 산출하였다.
NMS 분석(Non-metric Multidimensional Scaling)을 이용하여 각 산지별 구상나무림의 종조성 분포를 2차원 상에 환경요인과 함께 배열하였으며, 통계검정 및 분석프로그램은 SPSS 17.0 (IBM), PC-Ord 6.0 (mjm software)를 이용하였다(McCune and Grace, 2002; McCune and Mefford, 2006). 분류된 군집별 환경특성을 분산분석을 실시하였고 사후분석(Post hoc)은 Turkey-test로 평균 간의 차이를 비교하였다. 종간 상대적 우세를 비교하기 위하여 Curtis and McIntosh (1951)의 상대우점치(Brower and Zar, 1977)를 분석하였다. 상대우점치는 (상대피도+상대빈도+상대밀도)/3으로 계산하였다. 종풍부도는 조사구 내 출현한 종수로 정의하였으며, 종다양성은 식생데이터를 기반으로 하여 Shannon-Weaver의 종다양도, 최대종다양도, 균재도를 분석하였다(Kent and Coker, 1992).
연구결과
토양이화학적 성질
구상나무림 각 산지별 토양이화학적 성질을 분석한 결과 Table 1과 같다. 토양의 화학적 성질을 덕유산, 지리산, 한라산의 순으로 비교해보면, pH는 4.07 ± 0.06, 4.09 ± 0.03, 4.42 ± 0.07로 나타나 덕유산에서 가장 낮은 pH로 나타났으며, 전기전도도는 0.54 ± 0.07 dS/m, 0.66 ± 0.07 dS/m, 1.06 ± 0.19 dS/m, 유기물함량은 104.21 ± 6.41 g/㎏, 135.30 ± 10.17 g/㎏, 139.00 ± 22.95 g/㎏으로 분석되었다. 치환성 양이온 및 양이온치환용량(CEC)의 경우 Al3+는 645.34 ± 75.17 ㎎/㎏, 412.92 ± 31.12 ㎎/㎏, 254.84 ± 48.15 ㎎/㎏,으로 나타났으며, Mg2+, K+, Na+은 한라산에서 가장 높은 수치로 나타났으며, Ca2+는 지리산에서 가장 높은 수치로 나타났다. CEC의 경우 한라산에서 가장 높은 수치로 분석되었다. 토양의 화학적 성질의 통계적 유의성을 확인하기 위하여 ANOVA 분석을 시행하였으며, 사후검정으로 tukey의 검정방법을 이용하였다. 통계분석 결과. 유기물함량과 Ca2+를 제외한 모든 요소에서 통계적 유의성이 인정되었다 (p < 0.05).
Table 1.
Soil physico-chemical properties and environmental factor of each community ( ± SD, ns indicates that the item did not show significant different differences among treatments at the 5% level, *: p < 0.05, **: p < 0.01)
토양의 물리적 성질을 덕유산, 지리산, 한라산의 순으로 분석해보면, 모래함량은 33.50 ± 0.09%, 41.90 ± 0.04%, 44.22 ± 0.07% 로 나타나 한라산에서 가장 높은 수치고 분석되었으며, 미사함량은 57.72 ± 0.05%, 49.81 ± 0.12%, 51.75 ± 0.05%로 나타났으며, 점토함량은 8.80 ± 3.00%, 8.26 ± 1.47%, 3.94 ± 4.56%로 나타났다. 석력비율은 36.15 ± 2.64%, 26.57 ± 1.38%, 25.89 ± 4.52%로 분석되었다. 토양의 물리적 성질에 대한 ANOVA 분석결과, 미사함량은 제외한 나머지 요소에서 유의적인 차이가 있는 것으로 분석되었다(p < 0.05).
경관지수 분석
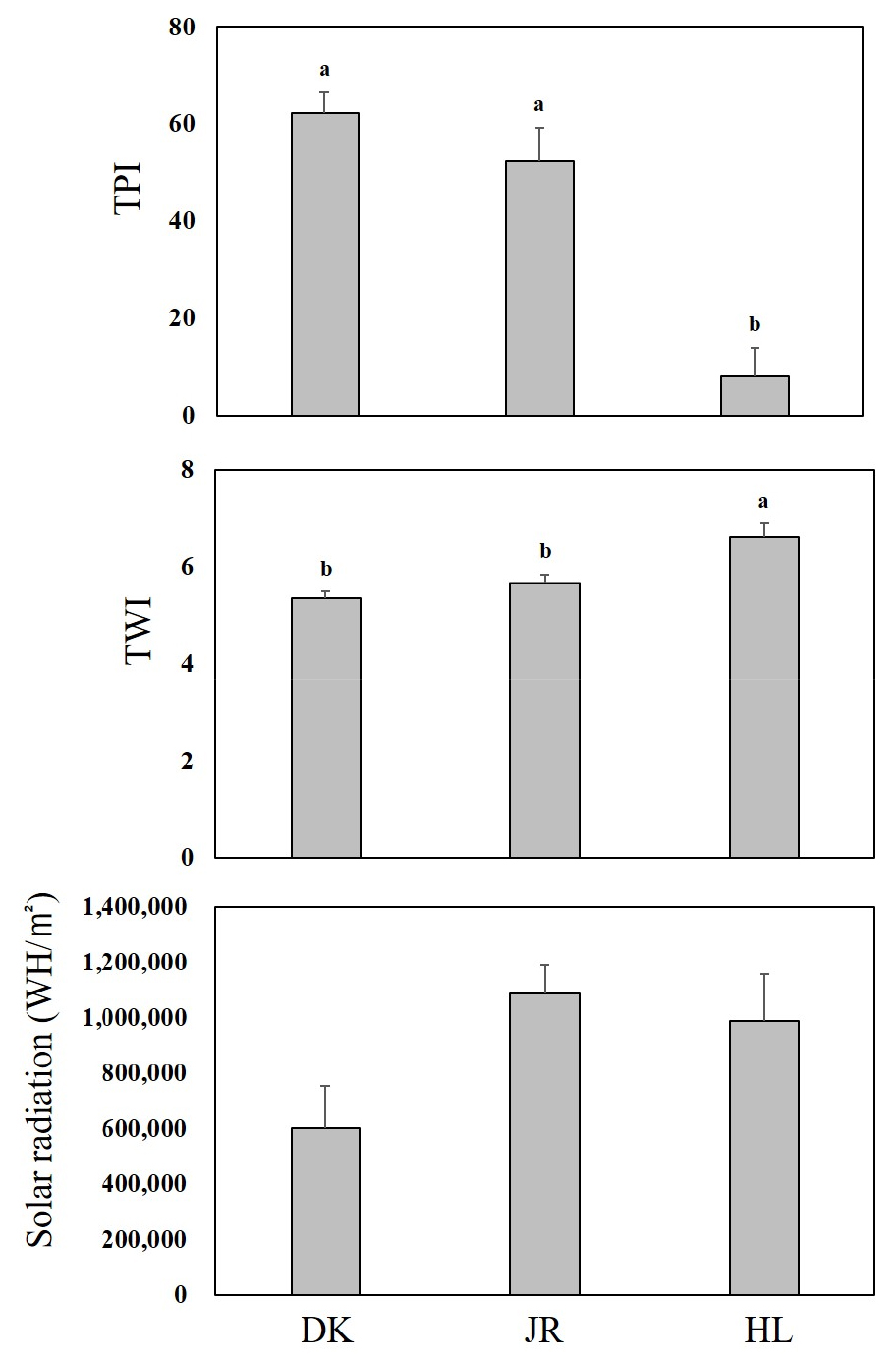
산지별 구상나무림의 경관지수를 분석한 결과 Fig. 3과 같다. 덕유산, 지리산, 한라산의 순으로 지형지수는 62.2 ± 4.5, 52.4 ± 6.9, 8.1 ± 5.8로 한라산에서 두 산지에 비하여 매우 적은 지형지수를 나타내었다. 수분지수의 경우 5.3 ± 0.1, 5.6 ± 0.2, 6.6 ± 0.3으로 나타났으며, Solar radiation은 602,406 WH/㎡, 1,086,677 WH/㎡, 990,267 WH/㎡로 분석되었다. 각 산지별 경관지수의 평균값의 차이는 지형지수와 수분지수에서는 유의적인 차이가 나타났으나(p < 0.05), Solar radiation 에서는 유의적인 차이가 나타나지 않았다(p > 0.05).
종조성 분포
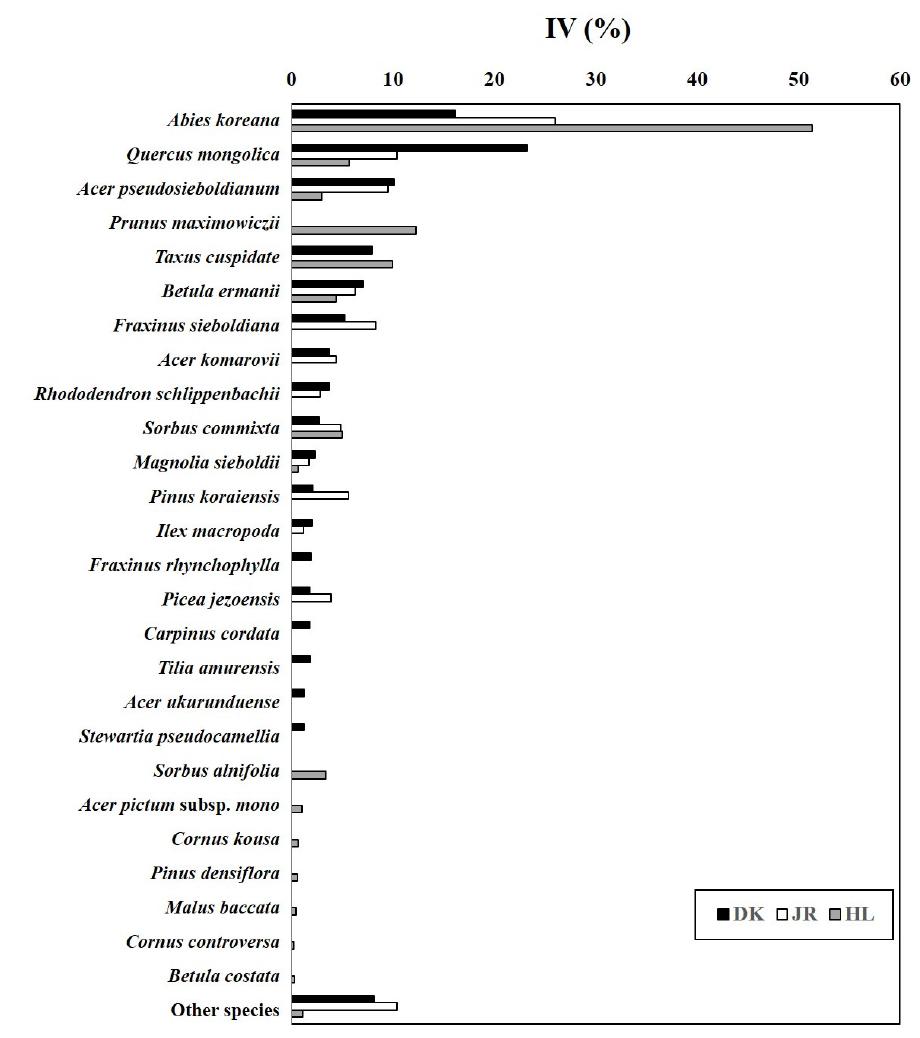
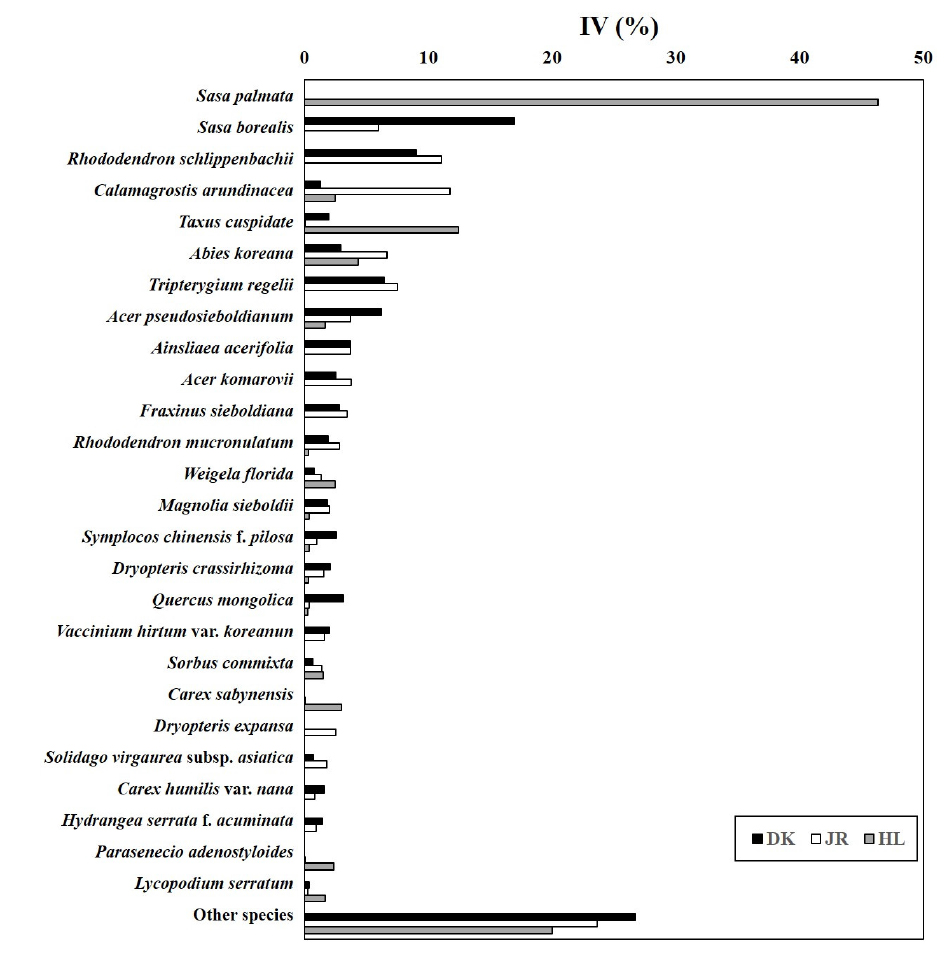
구상나무림의 목본수종의 중요치를 확인한 결과 Fig. 4와 같다. 덕유산의 경우 신갈나무(23.2) > 구상나무(16.1) > 당단풍나무 (10.1) > 주목 (8.0) > 사스래나무 (7.0) 등의 순으로 나타났으며, 지리산은 구상나무 (26.0) > 신갈나무 (10.4) > 당단풍나무 (9.5) > 쇠물푸레나무 (8.3) > 사스래나무 (6.3) 순으로 나타났다. 한라산의 경우 구상나무 (51.3) > 산개벚지나무 (12.3) > 주목 (10.0) > 신갈나무 (5.7) > 마가목 (5.0) 등의 순으로 나타났다. 국내 구상나무림의 대표산지에서도 각 수종의 우점정도는 상이한 것으로 나타났으며, 특히 구상나무의 분포가 가장 많은 곳은 한라산 > 지리산 > 덕유산의 순으로 나타났다. 덕유산의 경우 구상나무 임분보다 신갈나무의 중요치가 더욱 높게 나타났다.
하층식생의 중요치를 분석한 결과 Fig. 5와 같다. 덕유산은 조릿대 (16.9) > 철쭉 (9.1) > 미역줄나무 (6.4) > 당단풍나무 (6.2) > 단풍취 (3.7) 등의 순으로 나타났다. 지리산의 경우 철쭉 (11.1) > 실새풀 (11.7) > 미역줄나무 (7.5) > 구상나무 (6.7) > 조릿대 (6.0) 등의 순으로 나타났다. 한라산의 경우 제주조릿대 (46.3) > 주목 (12.4) > 구상나무 (4.3) > 실청사초 (3.0) > 붉은병꽃나무 (2.5) > 실새풀 (2.5) 등의 순으로 나타났다.
구상나무림의 하층식생과 상층식생의 각 산지별 종조성의 분포와 환경관의 관계를 살펴보기 위하여 NMS ordination을 통하여 2차원상에 배열하였다(Fig. 6).
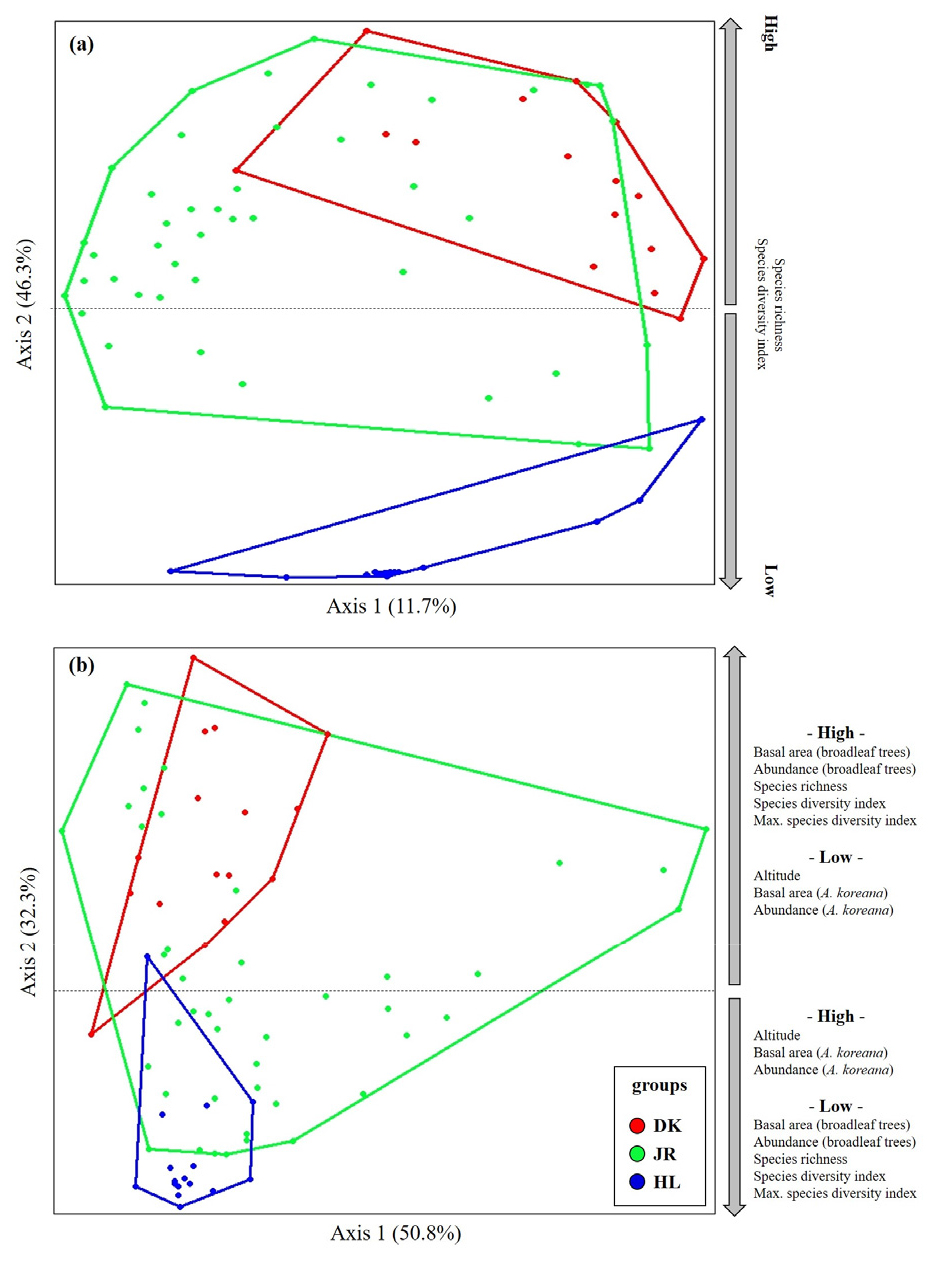
하층식생의 NMS ordination 결과 1축과 2축에서 가장 높은 설명력으로 나타났으며, 1축에서 11.7%, 2축에서 46.3%로 종합설명력 58.0%로 분석되었다. 하층식생의 종조성의 분포는 지리산에서 가장 넓은 공간으로 나타났으며, 한라산이 가장 좁은 종조성의 분포역을 나타내었다. 이는 하층식생의 중요치와 함께 살펴볼 때, 한라산의 경우 제주조릿대가 높은 비율로 차지하고 있어 타 식물종의 이입에 있어 공간적 경쟁에서 우점하여 나타난 결과로 판단된다. 또한 덕유산과 한라산에서 하층식생의 종조성의 분포가 서로 이질적으로 분석되었다. 이러한 하층식생의 종조성과 관계가 있는 인자는 종풍부도와 종다양성지수로 나타났다. 종다양성지수 및 종풍부도가 낮을수록 한라산 구상나무림의 종조성과 관계가 있는 것으로 분석되었다.
상층식생의 NMS ordination 결과 1축과 2축에서 가장 높은 설명력을 나타내었으며, 1축에서 50.8%, 2축에서 32.3%로 종합설명력 83.1%로 나타났다. 구상나무림 내 목본식생의 종조성 분포는 지리산이 가장 넓은 분포역으로 다양한 임분 구조를 지니고 있는 것으로 해석되었다. 또한 한라산 구상나무림의 경우 가장 좁은 종조성의 분포역으로 나타났다. 지리산의 경우는 타 산지에 비하여 구상나무의 생육지가 다양한 지형 및 생육환경에서 분포함을 의미하는 것이며 한라산의 경우는 분포역 내의 환경이 비슷하고 구상나무의 우점도가 상대적으로 높으며 다양성이 빈약하여 나타난 결과로 판단된다. 국내 구상나무림과 종분포의 환경인자간 상관관계는 활엽수의 흉고단면적, 활엽수 밀도, 하층식생 종풍부도, 하층식생 종다양성지수, 하층식생 최대종다양성지수, 해발고도, 구상나무의 흉고단면적, 구상나무의 밀도와 관련이 있는 것으로 분석되었다. 한라산 구상나무림의 경우 덕유산 구상나무림 보다 다소 높은 해발고도와의 상관관계가 있었으며, 낮은 활엽수 임분구조, 낮은 하층식생 종다양성과 연관이 있는 것으로 분석되었다. 국내 구상나무와 더불어 함께 서식하는 식물종의 분포는 지형, 토양 등 비생물적요인 영향보다는 활엽수의 흉고단면적, 종다양성 등과 같은 식생요인에 더 많은 영향을 받고 있는 것으로 나타났다.
흉고단면적 비교
산지별 흉고단면적을 비교한 결과 Fig. 7과 같다. 각 산지별 전체 상층식생의 흉고단면적과 구상나무 생육목 흉고단면적, 그리고 구상나무 고사목 흉고단면적을 비교하였다.
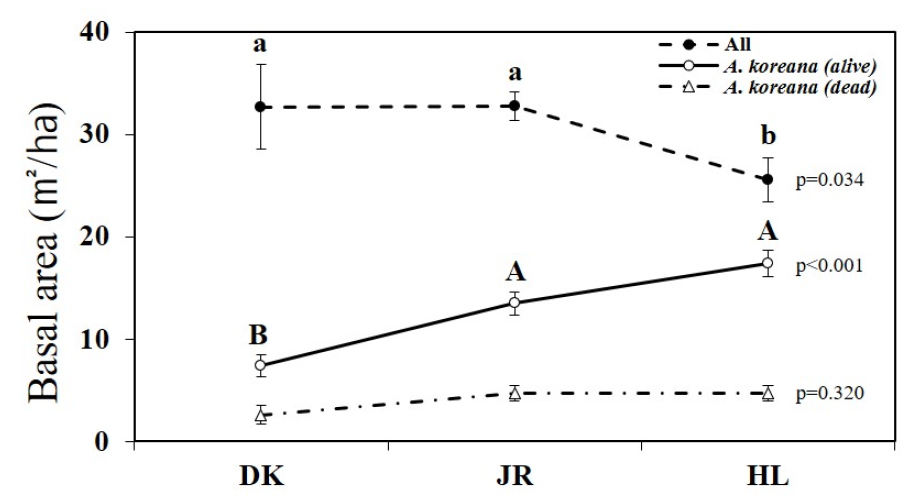
전체 흉고단면적의 경우, 덕유산 32.69 ± 4.17 ㎡/㏊, 지리산 32.77 ± 1.38 ㎡/㏊, 한라산 25.51 ± 2.16 ㎡/㏊로, 각 산지별 흉고단면적의 통계적 유의차가 있는 것으로 분석되었다(p < 0.05). 구상나무 생육목의 경우 덕유산 7.39 ± 1.10 ㎡/㏊, 지리산 13.48 ± 1.13 ㎡/㏊, 한라산 17.37 ± 1.32 ㎡/㏊로 나타나 산지별 유의적인 차이가 나타났다(p < 0.01). 구상나무 고사목의 경우 덕유산, 지리산, 한라산의 순으로 2.61 ± 0.94 ㎡/㏊, 4.70 ± 0.75 ㎡/㏊, 4.71 ± 0.78 ㎡/㏊로 나타났으나 통계적 유의차는 나타나지 않았다(p > 0.05).
흉고직경급 및 치수발생량
구상나무의 흉고직경급을 분석한 결과 Fig. 8과 같다. 구상나무 생육목의 경우 총 임목밀도의 경우 덕유산 183본/㏊, 지리산 281본/㏊, 한라산 644 본/㏊로 한라산에서 가장 많은 본수로 나타났다. 모든 산지에서 좌편향된 정규분포형의 흉고직경급으로 나타나 어린나무의 발생이 대체적으로 적은 임목밀도로 나타났다. 생육목의 수간형을 살펴보면 서있는 상태에서 생육(AS, Alived standing)가 덕유산 88.1%, 지리산 97.6%, 한라산 83.9%로 가장 많은 비율을 차지하였으며, 초두부가 부분고사된 채 생육하는 임목(AD, Alived deadtop)의 비율은 한라산이 11.1%로 가장 많이 나타났다.
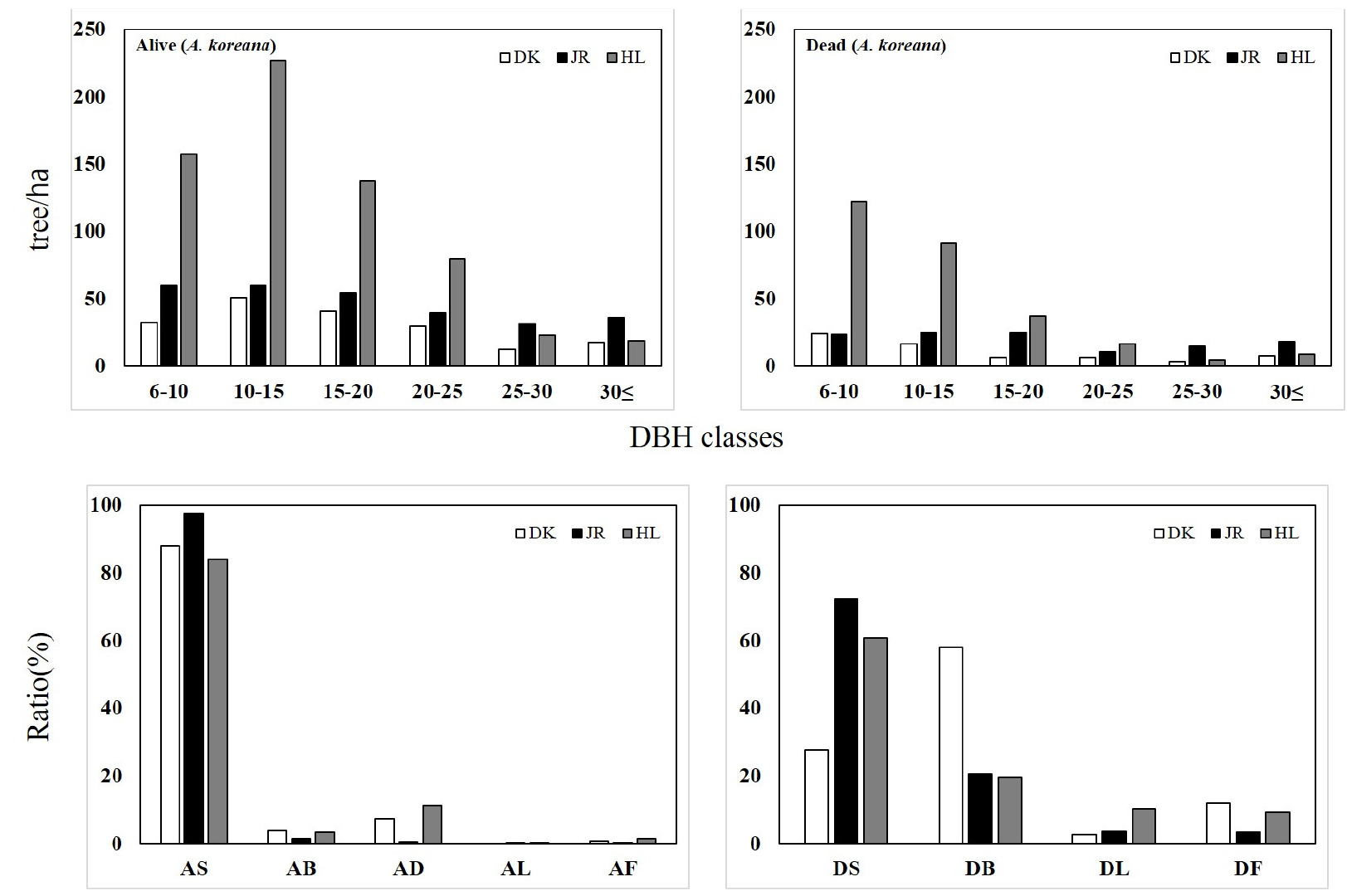
구상나무 고사목의 총 임목밀도는 덕유산 64 본/㏊, 지리산 116 본/㏊, 한라산 281 본/㏊로 한라산에서 가장 많은 고사목 임목밀도로 나타났다. 고사목의 수간형은 지리산과 한라산의 경우 서있는 채로 고사(DS, Dead standing)가 72.4%, 60.7%로 나타났으며, 덕유산의 경우에서는 부러진채 고사(DB, Dead broken)가 57.9%로 가장 많은 고사목 수간형으로 나타났다.
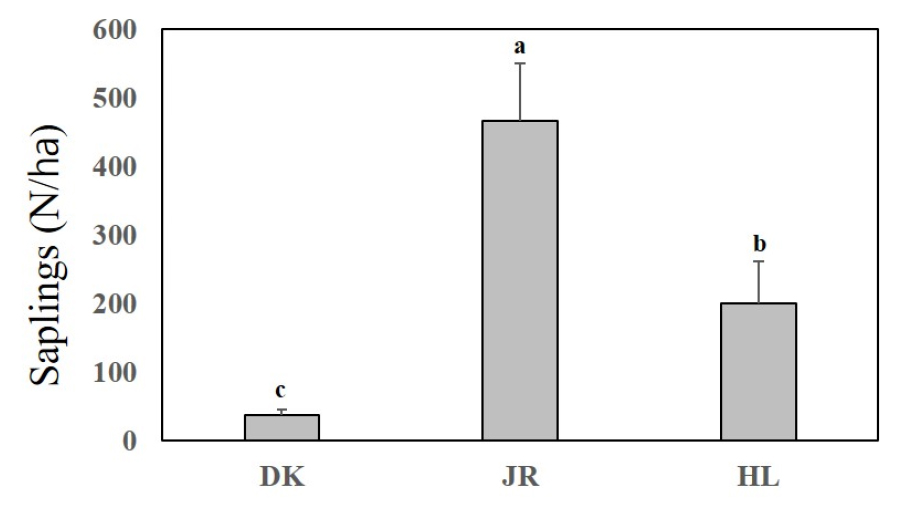
구상나무 치수발생량은 Fig. 9과 같다. 덕유산의 경우 39 ± 8 본㎡/㏊, 지리산 467 ± 83 본㎡/㏊, 한라산 200 ± 61 본㎡/㏊로 나타나 지리산에서 가장 많은 치수발생량으로 나타났다 세 산지의 구상나무 치수발생량은 통계적으로 유의한 차이가 있는 것으로 나타났다(p < 0.01).
상관관계분석
각 산지별 구상나무의 흉고단면적 및 임분밀도와 식생요인, 토양이화학적성질의 상관관계을 종합적으로 분석한 결과 Table 2와 같다. 흉고단면적과의 상관관계를 살펴보면, 생육목의 경우, 덕유산과는 어떤 환경인자간의 상관관계는 나타나지 않았으며, 지리산에서는 지형지수와 음의상관관계, 수분지수와 양의상관관계로 나타났으며, 한라산은 유기물, 나트륨, 칼슘 함량과 양의상관관계로 나타났다. 고사목의 경우 덕유산, 지리산 보다 한라산에서 더욱 환경인자에 민감한 것으로 분석되었는데, 해발고도, 지형지수, 전기전도도, 칼륨과 양의 상관관계로 나타났으며, 광량과는 음의 상관관계로 나타났다. 활엽수 경쟁과 구상나무 생육과의 상관관계를 살펴보면, 활엽수의 흉고단면적은 지리산의 치수발생량과의 양의 상관관계로 나타났으며, 활엽수 임분밀도는 지리산 구상나무 고사목의 임분밀도와 음의상관관계로 나타났다.
Table 2.
Correlation between basal area, density and environmental factors (pearson’s correlation method used, - and +: p < 0.05, -- and ++: p < 0.01)
임분밀도와의 상관관계를 살펴보면 덕유산은 지형지수와 전기전도도에서 양의 상관관계로 나타났으며, 지리산의 경우 지형지수와는 음의상관관계, 습윤지수와는 양의 상관관계로 나타났다. 한라산에서는 pH와 양의 상관관계로 나타났다. 고사목의 임목밀도는 주로 한라산에서 민감하게 나타났는데 해발고도, 지형지수, 전기전도도가 높을수록 고사목의 발생량이 많아지는 것으로 나타났다.
치수발생량의 경우 지리산에서는 통계적 유의성이 인정되었는데, 해발고도, 지형지수, pH에서 음의 상관관계로 나타났으며, 수분지수, 유기물함량, 알루미늄, 마그네슘, 칼슘, 석력함량과 양의 상관관계로 나타났다.
고 찰
본 연구는 국내 구상나무림을 대표하는 덕유산, 지리산, 한라산의 종조성과 환경인자간의 상관관계를 분석하여 향후 구상나무의 임분구조에 영향을 미치는 요인을 규명하는 기초자료 구축에 그 목적이 있다. 본 연구결과를 바탕으로 국내 구상나무림의 생육과 쇠퇴에 영향을 미치는 요인이 산지별로 상이하여 보전과 복원과 관련하여 환경에 맞는 적절한 관리방안을 도출해야 함을 판단하였다. 즉, 기 연구된 자료에 의하면 최근 구상나무의 쇠퇴원인에 대하여 수분스트레스 또는 풍해 등 여러 쇠퇴원인이 제시되어 왔지만 본 연구를 통해 개체군의 쇠퇴 및 유지 요인이 지역별로 다르게 적용되는 것으로 분석되었다.
토양이화학적 성질의 경우 동일한 구상나무 개체군을 조사하였으나, 덕유산과 지리산 그리고 한라산에서 유기물함량, 칼슘, 미사함량비를 제외한 나머지 요인에서 산지별 특이성이 있는 것으로 나타났다. 특히 구상나무의 임분밀도가 높고 흉고단면적이 높아 순림에 가까운 임분구조를 나타내었던 한라산 지역에서는 토양 양분이 풍부한 것으로 나타났으며, 모래의 함량이 타 지역보다 많고 점토함량이 적은 것으로 나타났다. 점토는 식물 내 보수력과 관련이 있으며, 모래함량이 높아 배수가 용이한 지형으로 분석된다. 이러한 결과로 볼 때 구상나무 임분발달이 가장 우수한 한라산 지역에서 토양 부족으로 인한 수분스트레스 영향은 적을 것으로 판단된다. 이러한 영향은 경관지수를 통해서도 알 수 있는데, 한라산의 수분지수가 높아 한라산 구상나무림에서는 수분스트레스가 임목발달 및 생육에 있어 큰 영향을 미치지는 않을 것으로 판단된다. 이는 한라산이 섬지형으로 이루어져 내륙에 위치한 지리산과 덕유산에 비하여 해양성 기후를 가지며, 이에 따라 연강수량이 많고 상대습도가 높기 때문에 나타난 결과로 판단된다(Park et al., 2009). 한라산의 구상나무 고사목과 solar radiation간의 음의 상관관계로 보아 광량이 낮을수록 고사목의 피도가 증가하였으며, 지형지수가 높은 지역, 즉 능선부로 갈수록 고사목의 피도가 높아졌다. 이로 보아 한라산 구상나무림은 광량과 바람에 의한 영향이 있을 것으로 판단된다. 반면 지리산에서는 경관지수에 의한 분석에서 지형지수가 낮고 수분지수가 높을수록 분포가 넓게 나타나는 것으로 나타나 수분에 의한 영향을 받고 있는 것으로 분석되었다. 치수발생량의 경우 지리산에서는 환경인자와 상관관계가 다수 나타나 치수발생이 상층목의 구상나무보다 더 민감한 것으로 나타났다.
본 연구 결과 덕유산의 경우 구상나무의 개체수 및 수관밀도가 매우 낮은 지역으로, 활엽수의 경쟁에 매우 많이 밀리고 있는 상황이다. 이는 Kim and Choo (2000)의 연구에서도 덕유산의 구상나무 임목밀도와 종조성이 매우 빈약하다고 언급한 것과 동일한 연구결과로 나타났다. 특히 신갈나무와 당단풍나무와 같은 Acer 속의 임분이 큰 중요치로 우점하고 있었다. 국내 신갈나무는 2차림(secondary forest)의 형태로 근주에서 맹아형태로 발달하여 향후 상층의 우점형태가 더욱 심화될 것으로 판단되어 덕유산 내 구상나무 개체군의 수관경쟁이 매우 불리해 질 것으로 판단된다. Acer 속의 식물은 높은 내음성과 내한성을 지닌 수종으로 치수 때 음수성을 가지는 전나무속(Abies spp)과 향후 층위경쟁으로 이어질 것으로 예상된다. ordination 결과 덕유산과 지리산의 종조성 분포역은 겹치는 부분이 많으나 한라산의 경우에는 이질적인 종조성을 가지고 있는 것으로 나타났다. 중요치 분석에서도 이러한 경향을 뒷받침 하고 있다. 이는 내륙에 위치한 지리산과 덕유산에 비하여 한라산의 현무암질의 토양조건과 해양성기후 등의 환경요인이 독특한 구상나무 임분 환경을 구성하는 것으로 판단된다. 또한 종조성의 이질성은 상층식생보다 하층식생에서 더욱 두드러지게 나타났는데, 이는 같은 환경조건의 변화 속에서 하층식생이 상층식생에 비하여 더욱 민감하게 반응하기 때문이다(Leith and Aston, 1961; Park et al., 2019). 상층식생의 경우 한라산의 구상나무 종조성은 높은 해발고도와의 상관성이 나타났으며, 덕유산의 구상나무림 종조성은 그 반대의 경향으로 나타났는데, 한라산 구상나무 임분구조가 덕유산보다 정량적으로 풍부하고 정성적으로 건강한 흉고직경급으로 나타나는 것으로 보아 구상나무 개체군은 높은 해발고도에서 안정된 임분으로 유지하고 있는 것으로 판단된다(Kim and Choo, 2000). 이러한 경향은 구상나무개체군이 더욱더 높은 해발고도를 양호한 생육지로 선호함을 의미하기도 하나, 한편으로는 지구환경변화에 따른 평균기온의 증가로 인하여 구상나무의 피난처(refugia)로써의 분포역이 점차 높은 해발고도로 이동하여 점차 분포역이 좁아지고 있음을 반증하기도 한다.
한라산의 하층식생의 종조성은 매우 낮은 종다양성지수로 나타났는데 이는 제주조릿대 군락이 매우 높은 밀도로 차지하고 있기 때문으로 판단된다(Kim, 1997). 하층식생의 발달은 Abies 속 치수의 생육공간을 차지하여 활착을 방해한다는 연구에 비추어 볼 때 제주조릿대가 우점한 곳의 구상나무 치수발달이 저해될 것으로 예상된다(Kim et al., 2019). 한라산의 구상나무는 낙엽층이 두꺼워 유기물층이 발달한 곳에 구상나무 성목과 치수의 발달이 많이 나타났다. 이는 21년전 Kim and Choo (2000)의 연구결과와는 상반된 분석으로 나타났는데, 임분구조의 연속성에 있어 치수발달은 낙엽층이 발달하지 않은 상황에서 공간적 우위를 차지하기 위하여 얕은 유기물층이 활착에 유리하나 향후, 치수 발달 및 생존에 있어서는 두터운 유기물층으로 인한 양호한 토양양분에 영향을 미칠 것으로 판단된다. 두터운 유기물 층은 식물체 성분과 생육에 필요한 질소함량을 증진시키며 소나무과 식물의 균근(Mycorrhiza)발생을 증진시킨다(Baier et al., 2006)
종합하면, 한라산의 구상나무 생육에 영향을 미치는 요인에 있어 수분스트레스가 큰 영향을 미치지 않은 것으로 분석되었으며, 지리산의 경우에서는 지형위치지수(TPI) 및 지형습윤지수(TWI)에 의해서 수분지수가 높은 곳에서의 구상나무의 분포가 많은 것으로 나타났다. 덕유산의 경우 능선부에 위치한 임분일수록 높은 임목밀도로 나타났다. 즉, 산지의 환경에 따라 구상나무의 분포에 영향을 미치는 요인이 일관적이지는 않은 것으로 나타났다. 현재까지의 구상나무림의 임분동태의 연구는 상층식생에 대한 연구가 주로 이루어졌으며, 임분유지와 갱신에 가장 큰 영향을 미칠 구상나무 치수의 생육발달에 대한 모니터링은 미흡하여 이에 대한 추가적인 모니터링 연구가 필요하다. 구상나무 임분유지 여부에 관한 연구중 치수발생량은 다소 단기적 모니터링에 의한 결과이므로, 이에 더 나아가 구상나무 치수의 연간 생장량을 조사하여 층위 발달에 대한 여부가 이루어지는지에 대한 연구자료 구축이 필요하다.
적 요
본 연구는 국내 대표적인 구상나무 자생지인 덕유산, 지리산, 한라산 대상, 종조성과 토양이화학적성질과 경관지수를 이용하여 구상나무 분포와 환경요인간의 상호관계를 분석하여 향후 구상나무 보전을 위한 기초자료를 마련하는 것이다. 표본점 조사구는 총 121개소를 조사하였다. 토양이화학적 성질의 경우 대부분 한라산에서 높은 수치로 기록되었으며, 지형지수는 덕유산에서 가장 높게 나타났고 습윤지수는 한라산, 광량은 지리산에서 가장 높게 나타났다. ordination 결과 덕유산과 한라산의 구상나무림 종조성이 매우 이질적인 종조성으로 나타났다. 구상나무의 흉고단면적의 경우 한라산에서 가장 높게 나타났으나, 활엽수 흉고단면적은 적어 한라산의 구상나무림이 육지에 있는 구상나무림 보다 순림형태로 나타났다. 치수발생량은 덕유산에서 매우 저조하여 향후 후계목의 수가 적어 점차 쇠퇴하고 있는 것으로 나타났으나 치수발생수 뿐만아니라 향후 치수의 생장량에 대한 모니터링이 지속적으로 진행되어야 할 것으로 판단된다. 환경인자와 구상나무 식생요인간의 상관관계를 살펴보면, 한라산 지역은 지형습윤지수와 큰 영향이 없었으며, 지리산 및 덕유산은 지형습윤지수에 영향을 미치는 것으로 나타났다. 또한 치수발생과 관련하여 지리산과 여러 환경인자간의 상관관계가 통계쩍 유의성이 있는 것으로 나타났다.
구상나무림의 쇠퇴와 유지와 관련하여 국내 학계에서 여러가지 가설이 제시되고 있으나 이에 대한 단기간의 조사를 바탕으로 한 분석이라는 점에서 연속적이고 여러 지형상황에 따른 모니터링이 필요할 것으로 판단된다. 또한 해양성기후인 한라산 지역과 대륙성기후를 나타내는 덕유산, 지리산 지역의 구상나무림의 쇠퇴 및 유지와 관련한 환경인자는 지역적 특성에 따라 차이가 있는 것으로 분석되었다.
