

서 언
재료 및 방법
식물 재료
종자 건조 및 저장
저장기간별 발아율 및 출아율 조사
육묘상에서의 생육 특성 조사
통계분석
결과 및 고찰
종자저장 3개월 후 발아 특성 조사
종자저장 15개월 후 발아 특성 조사
상토 파종 후 생육 특성
종자저장 27개월 후 발아 특성 조사
적 요
서 언
인삼(Panax ginseng C.A. Meyer)은 한국, 중국, 러시아 등 아시아의 극동 지역에서 자생하는 식물로, 뛰어난 약리 작용으로 오랫동안 중요한 약용식물로 사용되어 왔으며 고려 인삼의 인지도는 세계적으로 인정받고 있다(Lee et al., 2021; RDA, 2014).
인삼은 반음지 호냉성 다년생 식물로서 제한성장(determined growth)을 하고 생물적·비생물적 스트레스에 취약하다(RDA, 2014). 인삼은 종자로 번식하며 여름에 열매가 성숙하여 채종을 하지만 이때는 미성숙한 상태로 3 개월을 후숙시킨 후 일정기간 저온을 거쳐야 발아가 가능하다. 이는 인삼 종자가 형태생리학적 휴면(morphophysiological dormancy)을 가지고 있기 때문이며, 자갈과 모래 등과 함께 층적시켜 형태적 휴면타파를 위한 1차-후숙(warm-stratification)을 하면 배(embryo)가 형성되고 내과피가 열리게 되는데 이를 개갑(dehiscence)이라고 한다(Baskin and Baskin, 2014; Lee et al., 1986; Lee et al., 2016; Kwon et al., 1986; Kwon et al., 2001; Park et al., 2019). 이어 파종을 하거나 저온 저장을 하면 겨울 동안 저온을 거쳐 생리적 휴면타파를 위한 2차-후숙(cold-stratification)을 하여 봄에 발아를 할 수 있게 된다(Fig. 1).

종자 보급은 대부분 자가채종을 통해 이루어지며 시장에서 거래되기도 하는데, 이상기상 등으로 인해 종자 수급이 부족하거나 개갑이 불량하면 종자 가격에도 영향을 미치고 발아가 불량한 종자로 인해 피해를 보기도 한다. 본격적인 종자 생산은 3년생 이후에 가능하며 4년생 기준 개체당 약 50개의 종자만 형성이 되고 종자 형성은 뿌리의 생육을 감소시키므로 재배기간 동안 대부분 1회 정도만 채종한다(Kim et al., 2015b). 인삼 종자는 저장성이 낮고 대부분 당해연도에 수확해서 파종하기 때문에 저장에 대한 필요성이 높지 않아 뚜렷한 종자저장 기술이 알려져 있지 않지만, 갈수록 빈번해지는 이상기상의 피해로 인해 종자의 단기 또는 장기 저장 기술이 필요하다.
Lee et al.(2004)은 개갑이 완료된 종자를 건조시켜 5℃에서 6년간 저장이 가능하다고 하였으나 장기간 저장 시 지상부 출아가 불량하다고 하였다(Lee et al., 2004). Kim et al. (2015a)은 미개갑 종자를 건조시켜 -2℃에 1년, 2년, 3년, 5년간 종자를 저장하여 저장성을 조사하였는데, 저장 2년까지는 개갑과 발아가 양호하였지만 3년 후에는 개갑율과 발아율이 약 40%로 떨어졌다고 보고하였다. 두 연구 모두 건조 조건에 대한 조건이 명확하지 않으며, 노지에서의 출아율 확보에 대한 검증이 이루어지지 않은 문제점이 있다. 장기저장을 위해 초저온동결저장(cryopreservation)법이 개발되었지만, 이 방법은 유전자원을 보존하기 위해 소량의 종자를 고비용을 들여 저장하므로 대량 저장에는 적합하지 않다(Han et al., 2016).
종자는 건조와 저온 저장을 통해 저장성을 높일 수 있는데 이러한 종자를 저장성 종자(orthodox seed)로 분류하고, 건조나 저온에 취약하여서 수명이 짧은 종자를 난저장성 종자(recalcitrant seed)로 분류하며, 중간적 특성을 가진 종자들을 더 세분화하여 분류하기도 한다(Bonner, 1990; Ellis et al., 1990). 인삼은 난저장성 종자에 가깝지만, Lee et al.(2004)이 건조한 종자를 저온에 저장한 연구사례나 초저온동결건조 저장법에 관한 연구사례를 볼 때 건조나 저온 취약성에 대해 의문을 제기하게 되기도 한다(Han et al., 2016; Kim et al., 2008; Kim et al., 2015a).
인삼 종자의 장기 저장을 위해서는 최대 저장기간을 확보하는 방법을 선택하여야 할 것이나, 상황에 따라 종자 후숙 단계별로 적합한 방법이 필요할 것이다. 파종 후 남은 종자를 저장하거나 저장 후 즉각적으로 사용하기 위해서는 개갑이 완료된 종자를 저장하는 방법이 필요하다. 이에 본 연구는 개갑 인삼 종자의 저장 안정성을 구명하고자 종자의 수분함량이 저장성에 미치는 영향을 연구하였다.
재료 및 방법
식물 재료
시험재료는 2016년 7월 말~8월 초 충북 음성(batch #1, C)과 강원 진부(batch #2, G)에서 채취한 재래종 종자를 사용하였다. 채종 후에는 미개갑 상태이므로 인삼 종자 개갑방법(RDA, 2014)에 따라 종자망에 종자를 담고 모래와 자갈에 층적시키고 정기적으로 물을 뿌려주며 내과피가 벌어질 때까지 약 100일간 개갑처리를 하였다. 처리한 종자의 90% 이상 개갑되었고, batch #1과 batch #2의 100립중은 각각 5.7 g과 5.9 g이었다. 종자크기에 따른 선별은 따로 하지 않았으나 너무 크거나 작은 종자는 시험에 사용하지 않았다. 개갑이 완료된 종자를 흐르는 물에 씻은 후 물기를 제거하고 2℃에서 12월 중순까지 보관하다가 건조처리를 하였다.
종자 건조 및 저장
실험은 건조처리를 하지 않은 대조구와 내과피에 둘러싸인 진정종자(true seed)의 수분함량이 54%, 35%, 25%, 10%의 목표 수분함량(DMC = Desired Moisture Content)이 되도록 종자를 건조시킨 실험구로 구분하여 진행하였으며, 종자건조는 Rajametov et al.(2014)에 준하여 실시하였다. Batch #2의 경우 수분함량 45% 조건을 추가하였다. 차광막이 설치된 온실에서 선반 위에 망사를 깔고 그 위에 종자를 펼쳐 고르게 마를 수 있도록 뒤집어 주면서 건조하였으며, 대형선풍기(Hanil, Seoul, Korea)를 작동시켜 건조를 도왔다. 미리 무건조 종자의 중량과(SWi = Initial Seed Weight)과 수분함량(MCi = Initial Moisture Content)을 조사하였으며, 종자를 건조시키면서 진정종자의 중량이 DMC에 도달했을 때(SWDMC% = Seed Weight at DMC, 식1), 지퍼백(Cleanwrap, Kimhae, Korea)에 밀봉하여 담았다.
건조작업이 끝나고 수분함량을 50립씩 3반복으로 조사하여 DMC 여부를 확인한 후 -3.5℃, 상대습도 65.8% 저온 저장고(Seobin-go, Cheonan, Korea)에 저장하였다. 종자 batch와 건조 정도에 따른 종자의 최종 수분함량은 Table 1과 같다.
Table 1.
Seed moisture contents measured after seed dry at each DMC
저장기간별 발아율 및 출아율 조사
발아 특성은 저장 후 약 3 개월, 15 개월, 27 개월에 조사하였고, 이 시기는 일반적으로 농가에서 파종 후 발아시기인 3월말에 해당된다. 영하에 저장하였으므로 발아 전 온도변화에 적응시키기 위해 시험 3일 전에 2℃에 꺼내어 놓았다가 24시간 흐르는 물에 침지 후 다시 2℃에 3~4일간 저장하였다. 치상 전에 70% 에탄올로 1 분간 처리 후, 1% NaOCl을 10 분간 처리하여 표면 살균을 하고 수돗물로 냄새가 나지 않을 때까지 충분히 씻어주었다. Petri-dish에 여과지(Whatmann No. 2) 2 매를 깔고 멸균수를 넣어 각각 50립씩 3 반복으로 치상하고, 인삼 종자의 발아 적정온도인 10℃에서 배양하였다(Kwon et al., 1986; Lee et al., 2004). 1 주일 간격으로 발아율을 조사하였는데, 유근이 1 ㎜ 이상 출현하였을 때를 ‘유근발아’로 간주하고 유근발아율(GP = Germination Percentage), 평균발아일수(MGT = Mean Germination Time) 및 발아균일지수(GPI = Germination Power Index)를 산출하였다. MGT는 MGT = Σ(ti × ni)/N의 식을 이용하였으며, 이때 ti는 치상 후 조사일수, ni는 조사일 출아수, N은 총출아수이며 GPI는 GPI = GP/MGT × 100의 공식으로 구하였다(Choi et al., 2006; Lee et al., 2016). 발아 시작 약 3주 후에는 지상부가 종자 밖으로 나오기 시작하였는데 줄기와 어린 잎 조직이 종자 밖으로 완전히 나오면 ‘지상부 출아’로 판단하고(Fig. 1C) 눈으로 관찰하여 부패가 확실하거나 눌렀을 때 부패물이 나올 경우 ‘부패’로 간주하고 발생 개수 조사 후에 제거하였다. 지상부 출아수와 부패 종자수로부터 지상부 출아율(shoot emergence rate)과 부패율(seed rot rate)을 구하였다. 유근발아율, 지상부 출아율, 부패율은 치상 종자수 대비 각각의 백분율을 구하였는데 미개갑 종자가 있을 경우 이를 보정하였다.
육묘상에서의 생육 특성 조사
종자저장 후 2차 조사시기, 즉 15개월 차에 육묘 상자에 상토(피트모스 : 펄라이트 = 7 : 3)를 담고 150립씩 3 반복으로 파종하였다. 90% 차광막을 덮은 비닐하우스에서 재배하면서 출아율과 모잘록 발생률 조사하였으며, 약 7개월 후 반복 당 15개체의 근중을 조사하고, 3 반복의 평균을 구하였다.
통계분석
통계분석을 위하여 SAS v9.2 (SAS Institute Inc., Cary, NC, USA)을 이용하여 ANOVA 분석(Analysis of variance)을 수행하였으며, DMRT (Duncan’s Multiple Range Test) 5% 수준에서 일원 분산분석을 하여 유의성을 검증하였다(p < 0.05).
결과 및 고찰
종자저장 3개월 후 발아 특성 조사
2017년 3월, 저장 3개월 후 batch #1의 무건조 종자(C-Wet)의 유근발아율은 28.6%였으며, batch #2의 무건조 종자(G-Wet)의 경우 97.0%로 batch간에 차이가 있었다. Batch #1 종자의 건조도에 따른 유근발아율은 발아가 높은 순으로 C-DMC54, C-DMC35, C-Wet, C-DMC25, C-DMC10였으며 각각 69.6%, 38.7% 28.6%, 6.5%, 1.8% 발아하여 건조도가 높을수록 발아가 불량하였다(Table 2). 지상부 출아율은 유근발아율보다는 조금 낮았지만 건조도에 따른 차이는 비슷한 경향을 보였다. Batch #2는 전 처리구에서 유근발아율이 높았고 처리 종자 건조도에 따른 차이가 크지 않았지만, 지상부의 경우 수분함량 35% 이상 종자들은 98.2% 이상 출아했지만, 25% 이하로 건조된 종자들은 지상부 출아율이 낮았다(Table 2).
Table 2.
Germination properties of batch #1 and batch #2 ginseng seeds after 3 month of cold-storage
Values are the means ± standard error of the triplicate experiments. zRoot: root germination percentage, yEmergence: shoot emergence rate, xRot: seed rot rate, wMGT: mean germination time, vGPI: germination power index, uWet: seeds without drying, tDMC: seeds dried at determined moisture contents. sMeans with different letters within a column within a batch are significant at p < 0.05 by Duncan’s Multiple Range Test (DMRT).
MGT도 batch #1과 batch #2간에 차이가 났는데 batch #2의 MGT가 더 낮았으며, 종자 수분함량이 높을 때(> 35%) 더 낮은 경향이었다. GPI는 두 batch 모두 DMC54에서 높았으며, 25% 이하 건조 시 낮아졌다.
저장 전에는 휴면으로 인하여 발아력을 알 수 없으므로 종자의 생리적 휴면타파가 종료될 시점에서 종자의 초기 발아력을 조사하였다. 그러나 저온 휴면타파 기간 중 온도가 너무 낮으면 휴면타파가 지연될 수도 있는데(Suh et al., 2019), batch #1은 휴면타파가 지연되어 발아율이 낮은 것으로 보이며 batch #2는 휴면타파가 완전히 완료되어 두 batch 간 저온감응정도에 차이가 있는 것으로 보인다. 저장 6개월 후에 간이로 테트라졸리움 종자 활력검사를 하였을 때 일부 무건조 종자는 활력을 잃었지만 다수는 정상이었으며, 추가로 저온 처리 시 발아율이 증가하였다(data not presented). Batch #1이 상대적으로 낮은 발아율을 보인 것은 휴면타파가 완전히 종료되지 않은 것이 원인이었던 것으로 보이고, 무건조 종자는 종자활력과 휴면의 영향을 모두 받았을 것으로 생각된다.
종자저장 15개월 후 발아 특성 조사
2018년 3월, 저장 15개월 뒤 발아 특성을 조사하였을 때 batch #1의 유근발아율은 무건조 종자를 제외하고 1년 전보다 증가하여 C-DMC54, C-DMC35, C-DMC25는 각각 83.0%, 84.4%, 78.3% 발아하였고, C-DMC10과 C-Wet은 각각 7.3% 또는 12.0% 발아하였다(Table 3). 이후 C-DMC54와 C-DMC35의 지상부는 각각 72.9%, 52.0% 출아하여 출아율이 1년 전보다 높아졌지만, C-DMC25와 C-DMC10은 역시 출아하지 못하였고, C-Wet은 종자 부패가 54.9% 발생하여 발아율이 낮았다. 발아 기간 중에 부패가 나타난 것은 저장 기간 중에 이미 활력을 잃은 것으로 판단된다.
Table 3.
Germination properties of batch #1 and batch #2 seeds after 15 month of cold-storage
Values are the means ± standard error of the triplicate experiments. zRoot: root germination percentage, yEmergence: shoot emergence rate, xRot: seed rot rate, wMGT: mean germination time, vGPI: germination power index, uWet: seeds without drying, tDMC: seeds dried at determined moisture contents. sMeans with different letters within a column within a batch are significant at p < 0.05 by Duncan’s Multiple Range Test (DMRT).
Batch #2의 G-DMC54, G-DMC45, G-DMC35는 모두 93% 이상 유근이 발아하였으며, G-DMC25, G-Wet, G-DMC10이 각각 53.5%, 42.9%, 30% 발아하여, 무건조 종자와 수분함량 25% 이하 종자들은 1년 전보다 유근발아율이 낮아졌다(Table 3). G-DMC54와 G-DMC45의 지상부 출아율은 각각 97.6%, 98.2%으로 1년 전과 차이가 없었지만, G-DMC35는 그보다 낮은 79.6% 출아하였다. G-DMC25와 G-DMC10의 지상부 출아율은 더 낮아졌고, G-Wet과 G-DMC25는 부패가 많이 발생하였다. G-DMC25의 부패율이 높았지만, 3 반복 중 한 반복에서만 부패가 심하게 발생하였던 것으로 C-DMC25의 경우 부패율이 낮았던 것으로 볼 때(Table 2), 이는 시험과정 중에 예외적으로 발생한 결과로 판단된다.
Batch #2의 G-DMC54와 G-DMC45의 MGT와 GPI는 전년도에 비해 큰 차이를 보이지 않았지만(Table 3), 그 외 처리구들의 MGT는 1년 전보다 더 높았고 GPI는 감소하였다.
두 batch 모두 무건조 종자들은 15개월 저장 후 부패가 많이 발생하여 저장 기간 중에 죽은 것으로 보인다. 또한 수분함량 25% 이하 종자들은 유근의 발아가 양호하여도 지상부 출아는 정상적으로 이루어지지 않았으나, 부패는 잘 나타나지 않았다. 지방 함량이 높은 종자는 건조하여도 동결에 민감한데 인삼도 배유 성분중 60% 이상이 지질로서 낮은 수분함량에도 수분 동결이 쉽게 와서 동해를 입기 쉽다고 한다(Kim et al., 2008; Yoon et al., 2005). 현재로선 유근의 발아율과 지상부의 출아율 차이를 설명하기 어렵지만 본 연구결과를 토대로 볼 때 수분함량 25% 이하로 건조하는 것은 인삼 종자 저장 조건으로 부적합해 보인다.
상토 파종 후 생육 특성
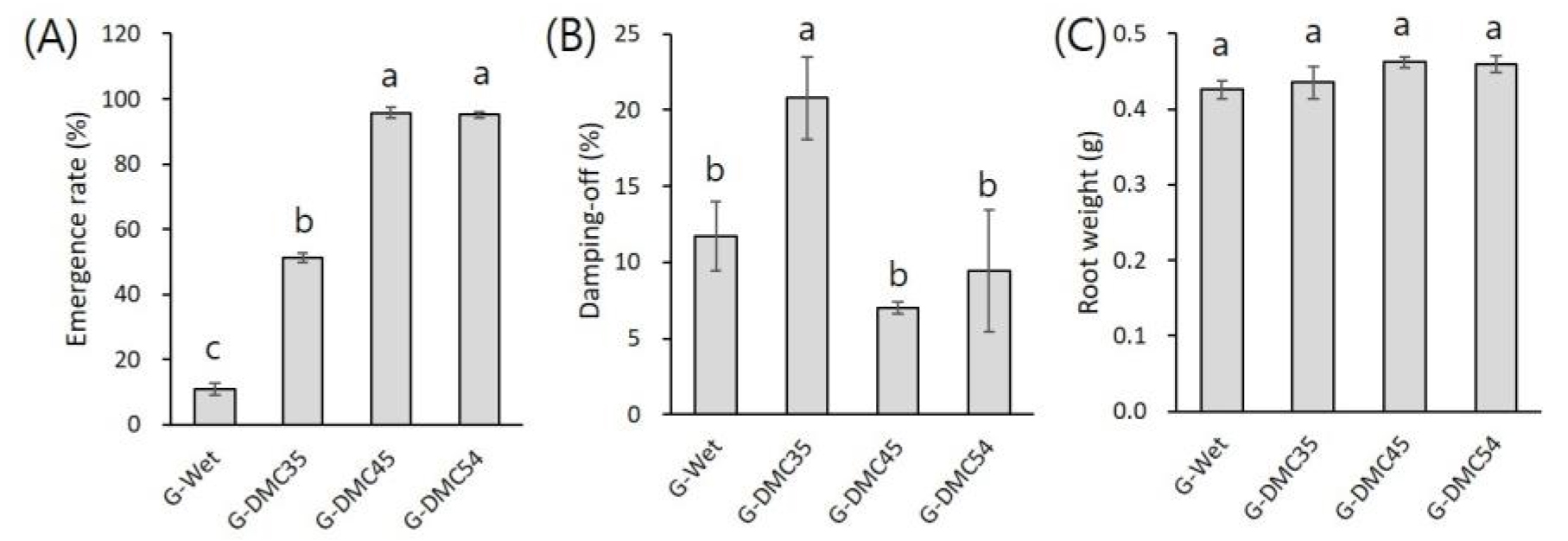
저장 15개월 후에 종자 활성이 더 양호한 batch #2 종자들을 상토에 파종하여 생육 특성을 조사하였다. G-DMC54와 G-DMC45는 각각 95.1%, 95.8% 출아하였고 G-DMC35는 51.3%, G-Wet은 11.1% 출아하여 기내 발아시험과 비슷한 경향을 보였다(Fig. 2). G-DMC35는 출아한 인삼 중 20.8%가 모잘록이 발생하여 생존수는 더 감소하였는데 이는 기내 실험으로는 확인할 수 없었던 것으로 종자 활력 검사시 노지 파종을 통한 검증 필요성을 보여준다. 근중은 G-Wet과 G-DMC35가 다른 처리구들에 비해 조금 작았지만 유의성은 없었다.
종자저장 27개월 후 발아 특성 조사
저장 27개월 후인 2019년도에는 2018년 조사에서 이미 발아력이 불량하였던 종자들은 제외하고 발아시험을 하였다. Batch #1의 C-DMC54와 batch #2의 G-DMC54는 각각 69.3%와 77.2% 유근이 발아하였지만, 지상부 출아는 30% 미만으로 활력이 많이 감소하였음을 알 수 있었다. Batch #2의 G-DMC45는 유근발아율은 45.7%이었으나 지상부 출아율은 12.1%로서 G-DMC54보다 종자 활력이 낮았다. C-DMC35와 G-DMC35는 전혀 출아하지 못하고 부패하였다.
본 연구 결과 수분함량에 따라 인삼 종자의 저장성에 확연한 차이가 있었는데 최대 수분함량에서 약 10% 내외로 부분 건조(partial dehydration)한 조건에서(DMC54 또는 DMC45) 저장성이 높았다(Table 3). 참나무과(Fagaceae) 종자들은 난저장성 종자들로서 건조하면 발아력을 잃지만, 이 경우도 종자 수분함량을 약간 낮추어 완전히 밀폐되지 않은 용기에 저장할 경우 저장성을 높일 수 있다고 한다(Kim et al., 2012). 밤나무(Castanea crenata) 종자는 일반적으로 6개월 정도 저장이 가능하나, 종자 수분함량을 40~45% 상태에서 지퍼백에 넣어 1~3℃에서 보관하면 최장 3.5년까지 연장이 가능하고, 참나무속(Quercus L.) 종자는 종자 수분 30~50% 상태에서 1~3℃ 저장 시 최대 3~4년까지 저장할 수 있다고 한다.
부분 건조 저장 방법은 저장성 종자 특성을 가진 일부 오리나무(Alnus glutinosa)나 자작나무(Betula pubescens) 등 온대 수목 종자들에서도 사용되어왔다. 이들은 휴면을 가지고 있어서 수분이 있는 조건에서 저온으로 휴면타파를 시켜야 하는데 휴면타파가 완료된 종자들이 파종 시기가 지연될 경우 사전 발아(pre-germination)를 하여 문제가 될 수 있다. 이때 휴면타파 기간 동안 최대 수분함량보다 조금 낮게 종자 수분을 유지함으로써 발아를 억제하면서도 종자 활력을 장기간 유지할 수 있다고 한다(Derkx, 2000; Gosling et al., 2003; Jones and Gosling, 1994; O’Reilly and Atrip, 2007).
부분 건조 조건과 수분함량에 큰 차이가 없는 무건조 종자들은 저장 시간이 길어질수록 활력을 상실하였는데(Table 3, Table 4), 이는 0℃ 이하에서 동결 피해를 받았기 때문으로 생각된다. 0℃ 이상에서 저장 할 경우는 휴면이 종료되면 발아를 해버리는 문제가 있다(Suh et al., 2017). 이에 비해 부분 건조 종자들이 저장 기간을 1년 정도 연장할 수 있었던 것은 건조 과정을 통해 저온에 대한 저항성이 증가하였기 때문이라고 생각된다. Kleinwächter et al. (2014)에 따르면 종자의 건조 내성은 dehydrin에 의해 증가하며, 심지어 이례적인 난저장성 종자들에서도 적용이 된다고 하였는데 이것에는 참나무속의 떡갈나무(Q. robur)도 포함된다. 고려 인삼에는 9개의 고유한 dehydrin 유전자(PgDhn)가 확인되었는데, 많은 경우 이 유전자의 발현은 저온 및 건조조건에서 증가하며, PgDhn9의 경우 종자에서 특이적으로 발현된다(Ha et al., 2006). 또한 인삼 종자의 초기 동결온도(initial freezing temperature)는 종자의 수분함량이 낮을수록 낮아진다고 하였다(Kim et al., 2008; Lee and Proctor, 1996). 이러한 물리·생물학적 기전이 건조 및 동결에 대한 내성을 부여하였는지는 실험을 통해 밝혀져야 할 것이다.
Table 4.
Germination properties of batch #1 and batch #2 seeds after 27 month of cold-storage
Values are the means ± standard error of the triplicate experiments. zRoot: root germination percentage, yEmergence: shoot emergence rate, xRot: seed rot rate, wMGT: mean germination time, vGPI: germination power index, uDMC: seeds dried at determined moisture contents, tWet: seeds without drying. sMeans with different letters within a column within a batch are significant at p < 0.05 by Duncan’s Multiple Range Test (DMRT).
종자가 건조될수록 활력이 감소하지만 부분 건조 조건에서 저장성이 좋아진다는 점에서 참나무과 종자들과 유사하므로 인삼 종자를 온대성 난저장성 종자로 분류하여도 좋을 것 같다. 또한 저장온도가 0℃에 가까운 온도에 저장이 가능하다는 점도 공통적인 특징이다(Bonnor, 1990; Kleinwächter et al., 2014). 참나무과 종자들도 최적 저장온도는 다르지만 서로 0℃에 가까운 온도, 즉 슈마드 참나무(Q. shumardii Buckl.) 종자는 -1.5℃에서 가장 잘 생존하고, 흑참나무(Q. nigra L.) 종자는 3℃에서 더 잘 생존하며, 떡갈나무 종자의 경우 -2℃에서 더 잘 생존한다고 한다(Bonner, 1990; Connor and Bonner, 1999; Noland et al., 2013).
Batch #1의 경우 휴면타파가 batch #2보다 지연되는 경향이 있었는데 종자 활력도 더 빨리 감소하였다. Batch #1과 #2는 재배지가 다른 차이도 있지만, 재배 과정에서 batch #1은 batch #2보다 불량한 환경에서 재배되었는데 이로 인해 종자 건전도가 떨어져 저장성이 낮아진 것으로 보이므로, 종자의 장기 저장을 목표로 할 때에는 모본의 관리도 신경써야 할 것이다.
이 연구를 통해 부분 건조 방법으로 인삼 종자의 저장성을 1년간 연장할 수 있다는 결과를 얻었다. 그러나 1년 이상 저장성을 유지하기 위해서는 보완이 필요하다. 인삼은 참나무과 종자들과 달리 내과피가 두껍게 둘러싸고 있는데 건조된 내과피가 수분 흡수제로 작용하여 종자 수분함량이 초기 조건으로 유지되지 않는 것을 확인한 바 있다(data not presented). 이로 인해 저장성에도 영향을 받았을 것으로 추측되며, 이를 방지하기 위한 방안이 필요하다. 또한 본 연구에서는 -3.5℃에 종자를 저장하였는데, 이는 다른 참나무과 종자의 예보다 낮은 것으로(Connor and Bonner, 1999; Noland et al., 2013) 최적 저장 온도에 관한 연구도 필요하다. 기후변화로 인한 이상기온은 양질의 종자를 충분히 확보하는 데 문제가 될 수 있다. 본 연구 결과를 활용하면 종자를 어렵지 않게 대량으로 저장할 수 있으므로 변화하는 환경재난에 대응할 수 있기를 기대하는 바이다.
적 요
인삼은 다년생 식물로 종자 번식을 하는데, 수확 시 미성숙한 배(embryo)를 가지고 있어 후숙이 필요하다. 기후변화에 대비하고 안정적인 종자 보급을 위해서는 단기 및 장기간 대량으로 종자를 저장할 수 있는 기술이 필요하다. 본 연구에서는 2개 지역에서 얻은 개갑된 인삼 종자를 수분함량을 달리하여 -3.5℃에 보관하고, 저장 3, 15, 27개월 후에 유근과 지상부 출현을 관찰하여 발아력 검정을 통해 종자 저장성을 조사하였다. 3개월 후, 전반적으로 batch #1보다 batch #2에서 발아율이 높았고, 그리고 수분함량이 35% 이상에서만 발아가 원활하였다. 15개월 후, batch #2에서 수분함량이 45~54%인 부분 건조된 종자들에서 가장 높은 발아율을 보였고, 대부분의 무건조 종자는 부패하며 발아에 실패하였다. 수분함량 25% 이하로 건조된 종자들 역시 유근발아 또는 출아가 불량하였다. 부분 건조 종자들은 토양에서의 발아 및 생육도 우수하였지만, 1년 뒤에는 역시 활력을 유지하지 못하였다. 요약하면, 인삼 종자는 온대성 난저장 종자의 특성을 가지는 것으로 보이며, 부분 건조 조건에서는 종자의 수명을 연장할 수 있어 1년 정도의 단기 저장에 활용할 수 있을 것으로 판단된다.
