

Introduction
Materials and Methods
Reagents and antibodies
DKE preparation
Cell culture and differentiation
Cell viability and cytotoxicity assays
Oil Red O staining and measurement of the triglyceride content
Western blotting
Statistical analysis
Results
DKE inhibits adipocyte differentiation
DKE regulates adipogenesis- and lipogenesis-related proteins expression
DKE activates the AMPK signaling pathway
Discussion
Introduction
Obesity is caused by an imbalance between excessive energy intake and consumption and is characterized by increased fat and blood lipid levels (Jung and Choi, 2014; Singla et al., 2010). Lipid accumulation refers to the development of adipocytes from preadipocyte precursor cells, and an increase in the number and size of adipocytes (Ali et al., 2013). As obesity can cause various complications such as hypertension and type 2 diabetes, continuous management and treatment are essential. Anti-obesity foods and food ingredients can effectively reduce visceral fat mass and prevent obesity and lifestyle-related diseases (Bray and Greenway, 1999; Saito, 2005).
Interest in natural products derived from edible plants with AMPK-activating properties and fewer side effects to manage lifestyle-related diseases caused by an imbalance in energy homeostasis is increasing (Hardie, 2014; Lyons and Roche, 2018; Sayed et al., 2023). Natural products play a notable role in traditional medicine owing to their various health-promoting properties, as there is an inverse correlation between dietary phenolic compound intake and mortality from various diseases (Sanchez-Moreno et al., 1998)
AMP-activated protein kinase (AMPK) regulates energy homeostasis by modulating glucose and lipid metabolism to maintain energy levels (Hardie, 2014). Several studies have demonstrated that AMPK activation can suppress preadipocyte differentiation and is accompanied by the suppression of transcription factors, such as peroxisome proliferator- activated receptor (PPAR) γ, CCAAT/enhancer-binding protein (C/EBP) α, and sterol regulatory-element binding protein-1c (SREBP-1c), which are required for adipogenesis (Daval et al., 2006; Huang et al., 2011). Additionally, under metabolic stress conditions, including oxidative stress and excessive exercise, AMPK promotes mitochondrial energy production and regulates energy homeostasis (Herzig and Shaw, 2018). Furthermore, AMPK activation exerts beneficial effects, such as increased fatty acid β-oxidation, decreased fatty acid and triacylglycerol synthesis, increased glucose uptake, and thermogenic activation (Choi et al., 2024; Richter and Ruderman, 2009; Srivastava et al., 2012). Therefore, AMPK is considered a therapeutic target for the treatment of lipid- related diseases, including obesity (Zhou et al., 2001).
Ripe persimmon fruits and leaves have long been widely used as herbal medicines (George and Redpath, 2008; Hossain and Shahidi, 2023). However, immature (unripe) persimmons are difficult to eat because of their bitter taste and have not been recognized as food. To confirm that immature persimmons can be consumed as food, we conducted a safety evaluation and found no problems with consumption (Ham et al., 2020). Moreover, concentrated tannins present in unripe persimmons are known to improve lipid metabolism (Gorinstein et al., 1998), promote alcohol metabolism (Kim et al., 2001), have antibacterial (Farha et al., 2020) and antioxidant (Lee et al., 2001) effects, and are thus expected to have high potential for development as a functional food material (Butt et al., 2015). We previously reported the anti-obesity effects of immature persimmon ethanol extract by analyzing changes in body weight, visceral fat weight, blood biochemicals, and adipogenesis-related gene expression levels in a high-fat diet-induced obese mouse model (Yoon et al., 2023). However, the mechanism of inhibition of lipid accumulation by immature persimmons in adipocytes has not yet been studied. Therefore, in this study, we evaluated the effects of immature persimmons on lipid accumulation in 3T3-L1 cells.
Materials and Methods
Reagents and antibodies
Dulbecco’s modified Eagle medium (DMEM), bovine serum (BS), fetal bovine serum (FBS), and penicillin-streptomycin were obtained from Gibco (Grand Island, NY, USA). The (LDH) Cytotoxicity Detection Kit was purchased from Promega (Madison, WIS, USA). Antibodies specific to PPARγ, C/EBPα, SREBP-1c, adipose-fatty acid-binding protein aP2, and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies against AMPKα, phospho-AMPK (p-AMPK) αThr172, acetyl-CoA carboxylase (ACC), and phospho-ACC (p-ACC)ser79 were acquired from Cell Signaling Technology (Beverly, MA, USA). Unless otherwise stated, all other reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA).
DKE preparation
DKE extract was prepared using the method proposed by Yoon et al. (2023). Whole immature persimmons were collected, washed, ground, and freeze-dried. Freeze-dried immature persimmon powder was extracted using 50% ethanol (Woori Ethanol Supply Company, Busan, Republic of Korea) for 24 h. The resulting ethanol solution was filtered and the filtered extract was concentrated under reduced pressure to obtain the extract.
Cell culture and differentiation
Mouse-derived 3T3-L1 preadipocytes were purchased from American Type Culture Collection (ATCC). Cell culture was performed at 37℃ and 5% CO2 using Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% (v/v) bovine serum (BS) and 1% penicillin/streptomycin. The cells were maintained by subculturing when the bottom of the T-75 cell culture flask was approximately 70% full. To differentiate preadipocytes, 3T3-L1 preadipocytes were seeded in a 12-well cell culture plate (1 × 105 cells/well) in DMEM supplemented with 10% BS and 1% P/S and cultured for 48 h until confluence. The medium was changed once more in the pre-confluent state and the cells were cultured for 48 h. At the confluent state (day 0 of differentiation induction), the culture medium was replaced with differentiation-inducing medium [DMEM containing 10% fetal bovine serum (FBS), 1% P/S, 1 μM dexamethasone (DEXA), 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), and 10 ㎍/mL insulin (INS)], and differentiation was induced for 2 days. Two days after the induction of differentiation, the cell culture medium was replaced with DMEM containing 10% FBS, 1% P/S, and 10 ㎍/mL insulin for 2 days. The medium was then replaced with DMEM containing 10% FBS and 1% P/S, and differentiation was induced for 2 days. DKE was processed simultaneously with medium replacement and maintained throughout the differentiation induction period.
Cell viability and cytotoxicity assays
Cell viability and cytotoxicity were examined using the MTT and LDH assays, respectively. Cell viability was examined using the MTT solution 72 h after treatment with DKE. The MTT solution in each well was completely removed and DMSO (100 mL) was added. The dissolved formazan was measured using a microplate reader at a wavelength of 595 ㎚. The lactate dehydrogenase (LDH) assay was performed using an LDH Cytotoxicity Detection Kit. The cells were treated with 1% Triton X 100 as a positive control to induce maximum LDH release, and absorbance was measured at 490 ㎚. Cell viability and cytotoxicity were calculated as the relative absorbance compared to that of the control.
Oil Red O staining and measurement of the triglyceride content
Staining was performed 7 days after simultaneous treatment with differentiation-inducing substances and DKE. Differentiation-induced 3T3-L1 cells were washed once with 1× PBS, fixed with 3.7% formalin for 1 h, and washed twice with distilled water. After completely removing the distilled water, Oil Red O staining was performed. Oil Red O solution was diluted with distilled water at a ratio of 6:4 and filtered through a syringe filter before use. After staining, the cells were washed thrice with distilled water and observed under an optical microscope. To quantify the accumulation of dyed lipids, Oil Red O was re-dissolved in isopropanol containing 4% NP-40 and the absorbance was measured at 520 ㎚.
Western blotting
3T3-L1 cells treated with DKE were washed 2 times with 1× phosphate-buffered saline (PBS) and harvested using lysis buffer supplemented with protease and phosphatase inhibitors. After centrifugation (15,000× g, 4℃, 20 min), the supernatant containing protein was obtained. Protein concentrations were determined using a Qubit4 fluorometer. 25-50 ㎍ of protein was electrophoresed on 8–12% mini gel SDS-PAGE and transferred to a polyvinylidene difluoride (PVDF) membrane. The PVDF membrane containing the transferred proteins was blocked with 5% skim milk or 5% BSA for 1 h and then incubated with the primary antibody. Subsequently, PPARγ antibody (1:500), C/EBPα antibody (1:2,000), SREBP-1c antibody (1:1,000), phospho-AMPK (1:2,000), ACC (1:1,000), and phospho-ACC (1:1,000) were incubated at 4℃ overnight, and A-FABP antibody (aP2; 1:5,000), AMPKα antibody (1:5,000), and β-actin antibody (1:10,000) were incubated for 1 h. After the primary antibody reaction, the PVDF membrane was washed with Tris-saline (TTBS) solution containing 0.1% Tween 20, and the peroxidase- conjugated secondary antibody was diluted to 1:5,000 or 1:10,000, reacted for 1 h, and then washed. Protein bands were visualized using an enhanced chemiluminescence (ECL) kit and a Vilber Western blot imaging system.
Statistical analysis
All results are presented as mean ± standard deviation (S.D.) of three independent experiments. The data were subjected to one-way analysis of variance, and statistical significance (p-value) was set at p<0.05 and p<0.01.
Results
DKE inhibits adipocyte differentiation
MTT and LDH assays were performed to determine the appropriate concentration of DKE for use in the cell experiments. When 3T3-L1 preadipocytes were treated with DKE at various concentrations (6.25, 12.5, 25, 50, 100, and 200 ㎍/mL), decreased cell viability and cytotoxicity were observed at concentrations of 100 and 200 ㎍/mL (Fig. 1A). Therefore, subsequent experiments were conducted using a maximum concentration of 50 ㎍/mL. To evaluate the effect of DKE on adipocyte differentiation, 3T3-L1 preadipocytes were induced to differentiate in the presence of MDI and DKE. Oil Red O staining on day 7 demonstrated a dose- dependent reduction in lipid accumulation in DKE-treated groups compared to the MDI-only control (Fig. 1B).
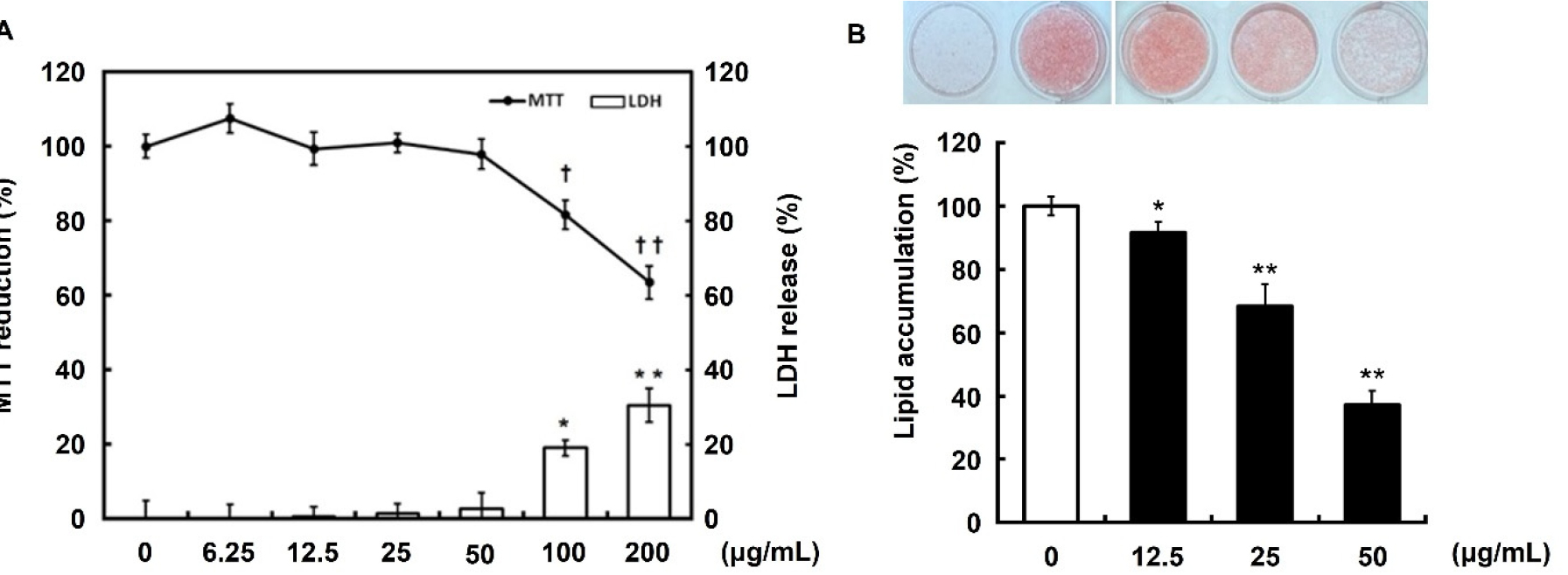
Fig. 1.
Effect of immature persimmon extract on cytotoxicity and lipid accumulation in 3T3-L1 cells. (A) Pre-confluent preadipocytes were incubated with the indicated concentrations of extracts for 72 h. Cell viability and cytotoxicity were measured using MTT and LDH assays, respectively. Data are presented as the mean ± S.D. of three independent experiments. *p<0.05, **p<0.01, †p<0.05 and ††p<0.01 compared to no-treated control. (B) 3T3-L1 cells were differentiated in MDI differentiation medium with different extract concentrations for 7 days. Differentiated adipocytes were stained with Oil Red O to visualize lipid accumulation. Lipid accumulation was assessed by quantifying the OD520. Data are presented as the mean ± S.D. of three independent experiments. *p<0.05 and **p<0.01 compared to no-treated control.
DKE regulates adipogenesis- and lipogenesis-related proteins expression
To further investigate the inhibitory effects of DKE on adipogenesis, we analyzed the protein expression of key adipogenic markers PPARγ, C/EBPα, and aP2 in MDI- induced 3T3-L1 cells treated with various concentrations of DKE (12.5, 25, and 50 ㎍/mL). Western blot analysis revealed a concentration-dependent suppression of these markers relative to the MDI-only group. Also, densitometric quantification confirmed significant decreases in PPARγ, C/EBPα, and aP2 protein levels, indicating a pronounced anti-adipogenic effect (Fig. 2A). Additionally, to assess the impact of DKE on lipogenesis, the expression of SREBP-1c and fatty acid synthase (FAS) was evaluated. MDI stimulation markedly upregulated SREBP-1c and FAS, whereas co-treatment with DKE significantly reduced their expression in a dose-dependent manner. Notably, SREBP-1c levels were almost completely abolished. These findings were further supported by quantitative analysis, which demonstrated significant reductions in both SREBP-1c and FAS protein levels (Fig. 2B).
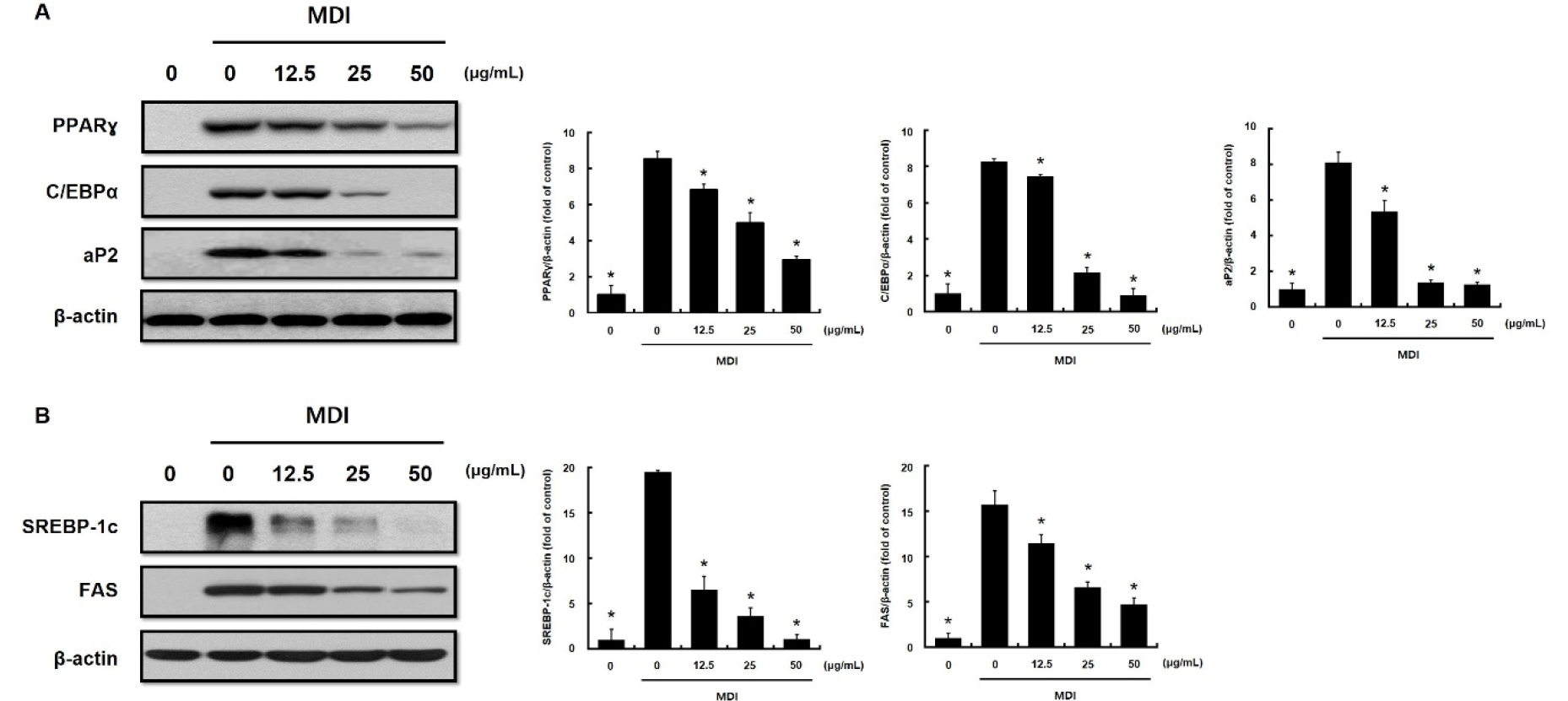
Fig. 2.
Effect of immature persimmon extract on the expression of adipogenesis-related proteins in 3T3-L1 cells. 3T3-L1 cells were differentiated in MDI differentiation medium with different extract concentrations, which was replaced every 2 days. Protein extracts were immunoblotted with specific antibodies that recognize (A) peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT/enhancer-binding protein α (C/EBPα), adipose-fatty acid-binding protein (aP2) and β-actin, (B) sterol regulatory-element binding protein-1c (SREBP-1c), fatty acid synthase (FAS) and β-actin. Data are presented as the mean ± S.D. of three independent experiments. *p<0.05 compared to MDI-only treated group.
DKE activates the AMPK signaling pathway
To elucidate the molecular mechanisms underlying the anti-adipogenic effects of DKE, we examined the activation of the AMPK signaling pathway. As shown in Fig. 3, DKE treatment led to a concentration-dependent increase in the phosphorylation of AMPK and its downstream target ACC. The ratios of p-AMPK/AMPK and p-ACC/ACC were significantly elevated at concentrations of 12.5, 25, and 50 ㎍/mL compared to MDI treatment alone. However, the total AMPK level remained constant, whereas the total ACC level slightly but significantly decreased at 25 and 50 ㎍/mL (Fig. 3). These results suggest that DKE suppresses lipid accumulation via AMPK activation and subsequent ACC phosphorylation.
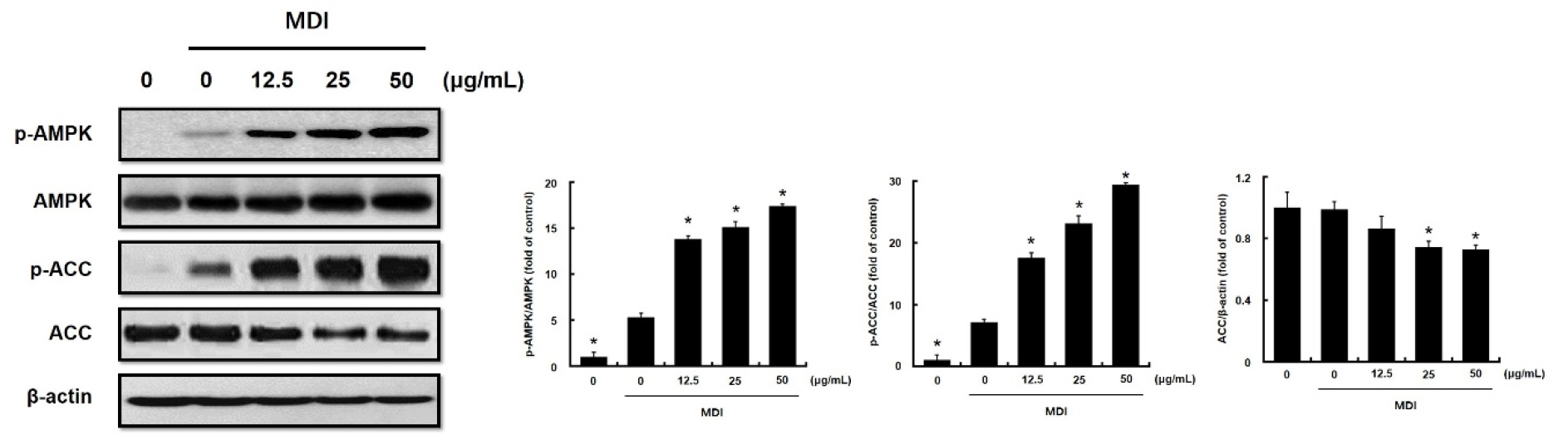
Fig. 3.
Effect of immature persimmon extract on AMPK and ACC phosphorylation in 3T3-L1 cells. 3T3-L1 cells were differentiated in MDI differentiation medium with different extract concentrations, which was replaced every 2 days. Protein extracts were immunoblotted with specific antibodies that recognize AMP-activated protein kinase (AMPK), phospho-AMPK, acetyl-CoA carboxylase (ACC), phospho-ACC and β-actin. Data are presented as the mean ± S.D. of three independent experiments. *p<0.05 compared to MDI-only treated group.
To confirm the involvement of AMPK in this pathway, Compound C, a selective AMPK inhibitor, was employed. Co-treatment with Compound C significantly attenuated the DKE-induced phosphorylation of AMPK and ACC, while increasing the expression of SREBP-1c, which had been downregulated by DKE treatment (Fig. 4). Quantitative analysis corroborated these results, showing that DKE- mediated increases in p-AMPK/AMPK and p-ACC/ACC ratios were significantly reversed by Compound C, while SREBP-1c and ACC expression levels were up-regulated. When CC and DKE were simultaneously treated, the phosphorylation of AMPK and ACC was restored, while the expression of SREBP-1c and ACC was decreased (Fig. 4). These findings collectively indicate that the anti-adipogenic and anti-lipogenic effects of DKE are mediated through activation of the AMPK signaling pathway.

Fig. 4.
Effect of immature persimmon extract on AMPK/ACC phosphorylation and SREBP-1c expression in 3T3-L1 cells preincubated with Compound C. 3T3-L1 cells were differentiated in MDI differentiation medium with different extract concentrations, which was replaced every 2 days. Protein extracts were immunoblotted with specific antibodies that recognize AMP-activated protein kinase (AMPK), phospho-AMPK, acetyl-CoA carboxylase (ACC), phospho-ACC, sterol regulatory- element binding protein-1c (SREBP-1c) and β-actin. Data are presented as the mean ± S.D. of three independent experiments. *p<0.05 compared to the Compound C-untreated group. #p<0.05 compared to the Compound C-treated group.
Discussion
Obesity is characterized by excessive fat accumulation, and inhibition of lipid storage in adipocytes is widely regarded as an effective strategy for its prevention (Fruh et al., 2021). In the present study, DKE significantly suppressed lipid accumulation in adipocytes, which was associated with a dose-dependent downregulation of key adipogenic transcription factors, including peroxisome proliferator-activated receptor γ (PPARγ), CCAAT/enhancer-binding protein α (C/EBPα), and adipocyte protein 2 (aP2) (Fig. 2A). These factors are well known to play central roles in the differentiation of preadipocytes into mature adipocytes (Huang et al., 2011). Furthermore, DKE treatment also led to a marked reduction in the expression of sterol regulatory element-binding protein-1c (SREBP-1c) and its downstream target fatty acid synthase (FAS) in a concentration-dependent manner (Fig. 2B). SREBP-1c is a lipogenic transcription factor that regulates lipid metabolism and homeostasis (Eberlé et al., 2004; Ferré and Foufelle, 2007). It is highly expressed in adipose tissue, where it promotes adipocyte differentiation by upregulating the expression of PPARγ and C/EBPα (Payne et al., 2009). The observed downregulation of SREBP-1c and its downstream effectors therefore provides further evidence that DKE suppresses adipogenesis at the molecular level. Collectively, these findings suggest that DKE exerts both anti-adipogenic and anti-lipogenic effects by attenuating the expression of key transcription factors involved in adipocyte differentiation and lipid biosynthesis. Furthermore, these results are in line with our previous in vivo study, which demonstrated the anti-obesity effects of DKE in a high-fat diet–induced obese mouse model, including reduced body weight gain and altered expression of fat metabolism–related genes (Yoon et al., 2023). Taken together, our data support the potential of DKE as a natural therapeutic agent for the prevention and management of obesity.
Persimmons are known for their diverse pharmacological activities, which are attributed to their strong antioxidant effects derived from various bioactive compounds, including tannins (Butt et al., 2015). Studies have suggested that tannins in persimmons may play a critical role in modulating various metabolic pathways, offering therapeutic potential for conditions such as obesity, diabetes, and cardiovascular diseases (Fukai et al., 2009; George and Redpath, 2008). Numerous studies have shown that tannins in persimmons can inhibit lipid droplet accumulation and stimulate lipolysis, thereby promoting the release of glycerol in adipocytes (Kim et al., 2008; Shin et al., 2014). Additionally, gallic acid produced by alkaline hydrolysis of tannins is believed to influence glucose metabolism by activating AMP-activated protein kinase (AMPK), a key regulator of energy homeostasis. AMPK activation improves glucose tolerance and decreases triglyceride levels in animal models of diet- induced obesity (Bak et al., 2013; Doan et al., 2015). Thus, it is suggested that the inhibitory effects of DKE on adipogenesis may be partly due to polyphenols, such as tannins, which are present in DKE. However, future studies are needed to identify the specific active compounds in DKE and investigate their effects to elucidate the mechanisms underlying its anti-obesity action.
AMPK plays a crucial role in regulating lipid metabolism by mediating the synthesis and breakdown of fatty acids (Srivastava et al., 2012). In this study, we identified the signaling pathway involved in AMPK phosphorylation by DKE. DKE did not alter AMPK protein levels, but did phosphorylate AMPK. In contrast, DKE decreased ACC protein expression, but increased ACC phosphorylation (Fig. 3). Activated AMPK has exercise-like effects on the human body, including the phosphorylation and inactivation of acetyl-CoA carboxylase (ACC), an enzyme required for ATP-consuming processes such as fatty acid synthesis (Richter and Ruderman, 2009). In addition, AMPK activation affects fatty acid oxidation by promoting ACC phosphorylation and lipid synthesis via regulation of SREBP-1c expression, thereby inhibiting ACC expression (Li et al., 2014). Therefore, the phosphorylation of ACC observed in Fig. 3 is likely a result of AMPK activation, and the decrease in ACC enzyme expression was caused by the inhibition of SRBPE-1c expression due to AMPK activation. These results are consistent with those of our previous study on the anti-obesity effects of DKE, which analyzed lipogenic gene expression levels in an obese mice fed with HFD (Yoon et al., 2023).
The selective AMPK inhibitor, Compound C, inhibits AMPK phosphorylation, thereby affecting the phosphorylation of the downstream factor ACC (Yoon et al., 2013). To determine whether the lipid accumulation caused by DKE was due to AMPK phosphorylation, we used Compound C. Compound C blocked AMPK phosphorylation, which was restored by DKE treatment (Fig. 4). In addition, AMPK activation was similar in CC-treated DKE and untreated DKE, suggesting that DKE is a potential AMPK activator. Interestingly, both Fig. 3 and 4 demonstrate that DKE affects AMPK activity. In addition, ACC phosphorylation, which is affected by AMPK phosphorylation, showed similar results to AMPK activation (Fig. 4). Previous studies by Ha et al. have shown that β-HIVS, a natural naphthoquinone compound, inhibits lipogenesis by activating AMPK, leading to downregulation of mature SREBP‑1c and lipogenic enzymes (Ha et al., 2016). Also, Berberine, a natural isoquinoline, activates AMPK and induces SREBP‑1c phosphorylation, thereby reducing lipogenic gene expression and intracellular fat accumulation (Jang et al., 2017). Another studies have shown that SREBP-1c expression and activation are regulated by AMPK signaling (Attal et al., 2022; Li et al., 2011). According to Fig. 4, it was found that SREBP-1c was decreased when AMPK was activated and then increased again when AMPK activity was inhibited, and followed ACC expression showed results similar to those of SREBP-1c expression. Therefore, it is assumed that the restoration of SREBP-1c expression is affected by the restoration of AMPK activation, and the inhibition of lipid accumulation caused by DKE treatment is due to their interaction.
Overall, this study showed that DKE exerted anti- adipogenic effects by downregulating adipogenic transcription factors via AMPK activation during adipogenesis. In conclusion, immature persimmons (Diospyros kaki Thunb.) may serve as a natural agent for the prevention and treatment of obesity via the AMPK pathway.
