서 언
재료 및 방법
실험재료
시료추출
세포배양
자가포식 측정
Transfection of small interference RNA (siRNA)
SDS-PAGE and Western blot analysis
통계분석
결과 및 고찰
작약 뿌리 추출물의 RAW264.7세포에서 자가포식 유도 및 p62/SQSTM1 생성에 미치는 영향
TLR 2와 4가 작약 추출물 매개 p62/SQSTM1 단백질 증가와 자가포식 유도에 미치는 영향
작약 추출물 매개 p62/SQSTM1 단백질 증가와 관련된 신호전달 분석
Nrf2가 작약 뿌리 추출물 매개 p62/SQSTM1 단백질의 증가에 미치는 영향
적 요
서 언
작약(Paeonia lactiflora)은 작약과(Paeoniaceae)의 다년생 초본식물로 한국, 중국, 시베리아 남동부 등 동북아시아 지역에서 한약재로 활용되고 있다(Lee et al., 2022). 작약은 간을 진정시키고 통증을 완화시키며, 혈액 영양 공급, 월경 조절 및 땀을 막는 데 널리 사용되어 왔다(Wang et al., 2022). 게다가 작약은 자가면역 질환, 류마티스 관절염 및 전신성 루푸스 등의 염증성 질환 치료에 널리 사용되고 있다(He and Dai, 2011; Zhang and Wei, 2020). 많은 연구를 통해 염증성 질환 치료제로의 작약에 대한 과학적 근거가 확보되고 있다(Chen et al., 2008; Kim and Ha, 2009; Li et al., 2007). 그러나 작약의 항염증 활성과는 반대로, 작약의 다당체들은 선천성 면역세포인 대식세포에서 면역 자극 인자의 분비를 촉진 시키고, 포식작용의 활성화를 유도한다고 보고되었다(Wang et al., 2022). 게다가, 작약은 후천성 면역세포인 T세포의 증식과 T세포의 분화를 촉진한다고 보고되었다(He and Dai, 2011). 대식세포와 같은 선천성 면역세포의 활성화는 후천성 면역 반응을 일으키는 데 중요하다고 알려져 있기 때문에(Geum et al., 2022; Quesniaux et al., 2010), 이러한 선행보고들은 작약은 선천성 면역 반응과 후천성 면역 반응을 동시 강화할 수 있다는 것을 보여주고 있다.
대식세포는 백신 유도 면역 반응에서 중요한 역할을 한다고 알려져서 대식세포의 활성화는 in vitro에서 신규 백신 보조제를 탐색하는데 중요한 지표로 활용되고 있다(Rimaniol et al., 2004; Quesniaux et al., 2010). 그리고 대식세포의 포식작용과 항원처리 능력의 활성화는 백신 보조제 개발의 첫 단계로 알려져 백신 보조제 개발을 위해 대식세포의 활성화를 유도하는 잠재적 소재의 탐색이 이루어지고 있다(Yamamoto and Naraparaju, 1998). 대식세포의 자가포식은 인체가 다양한 병원체에 대항하는 주요 면역기전 중 하나로 알려져 있고, 대식세포의 주요 기능인 식균작용, 전 염증성 사이토카인 방출, 염증 반응 및 항원 제시 기능에 관여한다(Wu and Lu, 2020). 게다가 대식세포에서 자가포식 유도는 대식세포의 항원 처리 과정 및 항원 제시기능을 강화해 선천성 면역 반응과 적응성 면역 반응 사이의 연결을 증가시킨다고 알려져 있다(Kirkegaard et al., 2004; Geum et al., 2022).
따라서, 본 연구에서는 작약이 대식세포의 활성화를 유도한다는 기존 연구를 바탕으로 작약이 대식세포의 자가포식에 미치는 영향과 그 작용기전 구명하였다.
재료 및 방법
실험재료
MTT, Griess 및 화학적 억제제(TAK-242, C29, PD98059, SB203580, SP600125, BAY 11-7082 및 LY294002)는 Sigma-Aldrich(St. Louis, MO, USA)에서 구입하였다. 1차 항체(p-PI3K, PI3K, Nrf2, p62/SQSTM1 및 β-actin), 그리고 2차 항체(Anti-rabbit)는 Cell Signaling(Bervely, MA, USA)에서 구입하였다.
시료추출
작약(Paeonia lactiflora) 뿌리는 경북 봉화에 위치는 봉화약용작물연구소에서 제공받았다. 40℃에서 열풍건조 후 파쇄된 작약 뿌리를 부피의 20배 증류수로 40℃에서 24시간 동안 추출하였다. 추출 후 원심분리하여 획득한 추출물을 동결 건조하였다. 동결건조 된 작약 뿌리 추출물(PLR)은 멸균수로 용해하여 실험에 사용하였다.
세포배양
American Type Culture Collection (Manassas, VA, USA)에서 구입한 마우스 대식세포인 RAW264.7 세포는 10% FBS, 100 U/mL penicillin과 100 ㎍/mL streptomycin이 포함된 DMEM/F-12 배지로 37℃, 5% CO2 하에서 배양되었다.
자가포식 측정
작약 뿌리 추출물의 RAW264.7세포에서 자가포식 유도는 Autophagy Assay Kit (Sigma-Aldrich)를 활용하여 측정하였다. RAW264.7세포를 96-well plate에 배양하여 세포가 well에 80% 이상 찼을 때, 작약 뿌리 추출물을 농도별로 24시간 동안 처리하였다. 작약 뿌리 추출물 처리 24시간 후, RAW264.7세포에 Autophagosome Detection Reagent를 첨가하여 37℃, 5% CO2에서 1시간 동안 배양하였다. 1시간 후 세포를 Wash Buffer로 4회 세척 하고, UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 형광광도(λex = 360/λem = 520 ㎚)를 측정하였다.
Transfection of small interference RNA (siRNA)
siRNA의 transfection은 TransIT-TKO transfection reagent (Mirus Corp., Madison, WI)를 활용하여 수행되었다. 96-well plate에 배양된 RAW264.7세포에 100 nM의 control-siRNA와 p62/SQSTM1-siRNA를 처리하고 48시간 동안 배양하였다. 48시간 후 작약 뿌리 추출물을 처리하여 RAW264.7세포의 자가포식 유도를 측정하였다.
SDS-PAGE and Western blot analysis
RAW264.7 세포를 1 × phosphate-buffered saline으로 2회 세척한 후, protease inhibitor cocktail (Sigma-Aldrich Co.)과 phosphatase inhibitor cocktail (Sigma-Aldrich Co.)이 포함된 radioimmunoprecipitation buffer (Boston Bio Products, Ashland, MA, USA)로 RAW264.7 세포로부터 단백질을 추출하였다. 추출된 단백질은 Bicinchoninic acid protein assay (Pierce Biotechnology Inc., Waltham, MA, USA)로 정량분석하였고, 동일량의 단백질을 SDS-acrylamide gel로 전기영동하고 PVDF membrane (Bio-Rad, Hercules, CA, USA)에 이동시킨 후 5% non-fat dry milk로 상온에서 1시간 동안 blocking 하였다. 1시간 후, 1차 항체를 5% non-fat dry milk에 용해시켜 4℃에서 16시간 동안 반응시킨 후 membrane을 0.05% tween-20이 포함된 tris-buffered saline (TBS-T)로 5분간 3회 세척하였다. 그 후 2차 항체는 5% non-fat dry milk에 용해시켜 membrane에 상온에서 1시간 처리하였고, TBS-T로 5분간 3회 세척 후 membrane은 ECL western blotting substrate (Amersham Biosciences Co., Little Chalfont, England)를 이용하여 단백질을 확인하였다.
통계분석
모든 결과는 3회 반복 측정 후 평균 ± 표준편차로 나타내었고, 처리간 유의성은 Student’s t-test로 검증하여 p-value 값이 0.05 미만일 때 통계적으로 유의하다고 판정하였다(Microsoft Exel 2010, Microsoft, Redmond, WA, USA).
결과 및 고찰
작약 뿌리 추출물의 RAW264.7세포에서 자가포식 유도 및 p62/SQSTM1 생성에 미치는 영향
작약 뿌리 추출물이 마우스 대식세포인 RAW264.7 세포에서 자가포식을 유도하는지 알아보기 위해, RAW264.7세포에 작약 뿌리 추출물을 처리하고 Autophagy Assay Kit로 자가포식 유도활성을 측정하였다. 그 결과, 작약 뿌리 추출물은 농도의존적으로 자가포식을 유도하는 것으로 나타났다(Fig. 1A).
자가포식은 리소솜(lysosomes)과 융합 된 자가포식체(autophagosome)를 통해 세포질 소기관과 단백질을 제거하는 시스템으로 알려져 있다(Delgado et al., 2009). 최근에 p62/SQSTM1은 유비퀴틴화된 단백질, 세포 내 병원체 및 손상된 미토콘드리아 등을 선택적으로 자가포식하는 데 중요한 역할을 한다고 보고되었다(Johansen and Lamark, 2011). p62/SQSTM1은 시퀘스토좀(sequestosomes)라고도 불리는 대규모 집합체/포함체(large aggregates/inclusions) 형성에서 중개자로 작용하며, 세포 내 병원체의 분해를 위해 자가포식체로 운반하는 기능을 한다(Into et al., 2012). 이러한 선행보고들은 p62/SQSTM1은 자가포식과정에서 세포 내 병원체 게거를 위한 자가포식체 형성의 플랫폼을 제공할 수 있다는 것을 의미한다. 따라서 우리는 작약 뿌리 추출물의 자가포식 유도에 p62/SQSTM1이 관여하는 알아보기 위해, 작약 뿌리 추출물이 농도별 또는 시간별로 처리된 RAW264.7세포에서 Western blot 분석으로 p62/SQSTM1 단백질의 수준을 조사하였다. 그 결과, 작약 뿌리 추출물이 처리된 RAW264.7세포에서 p62/SQSTM1의 단백질 증가가 나타났고, p62/SQSTM1의 단백질은 작약 뿌리 추출물 처리 6시간 째부터 급격한 증가가 확인되었다(Fig. 1B and 1C). 게다가 p62/SQSTM1 siRNA로 p62/SQSTM1이 knockdown 된 RAW264.7세포에서 작약 뿌리 추출물 매개 자가포식 유도가 감소하는 것으로 나타났다(Fig. 1D). 본 결과를 미루어 볼 때, 작약 뿌리 추출물은 p62/SQSTM1을 증가시켜 자가포식을 유도할 수 있는 것으로 판단된다.
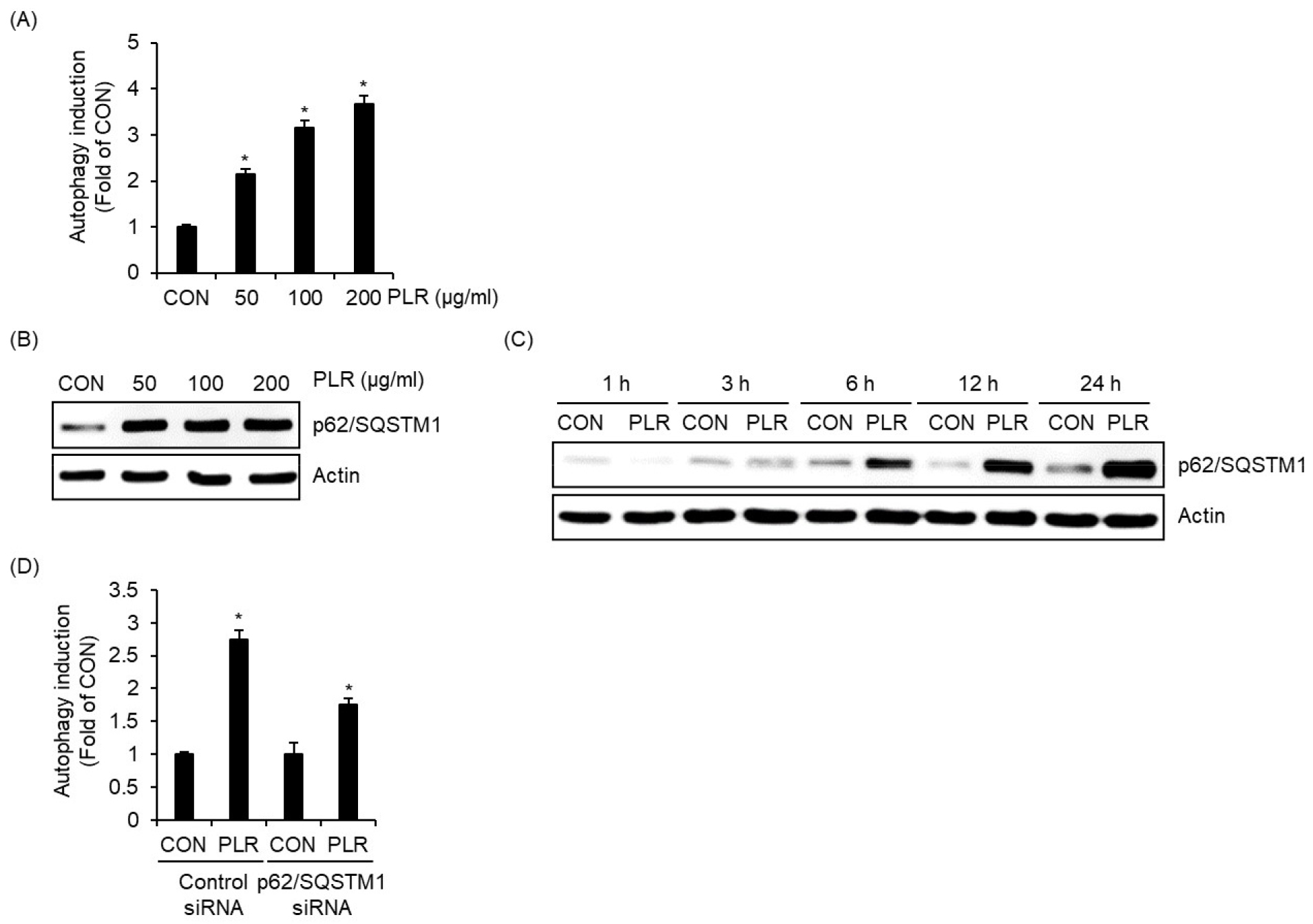
Fig. 1.
Effect of PLR on autophagy induction and p62/SQSTM1 level in RAW264.7 cells. (A) RAW264.7 cells were treated with PLR for 24 h. Autophagy induction was measured using Autophagy Assay Kit. (B) RAW264.7 cells were treated with PLR for 24 h. p62/SQSTM1 level was measured using Western blot analysis. (C) RAW264.7 cells were treated with PLR (50 ㎍/mL) for the indicatd times. p62/SQSTM1 level was measured using Western blot analysis. (D) RAW264.7 cells were transfected with control- or p62/SQSTM1 siRNA for 48 h, and then treated with PLR (50 ㎍/mL) for 24 h. Autophagy induction was measured using Autophagy Assay Kit.
TLR 2와 4가 작약 추출물 매개 p62/SQSTM1 단백질 증가와 자가포식 유도에 미치는 영향
대식세포는 TLRs를 통해 외부 병원체를 인식하여 체내 면역반응을 시작하는데, 이는 체내 면역방어에 매우 중요한 과정으로 알려져 있다(Chou et al., 2020). 최근 연구에서는 TLRs이 자가포식을 유도함으로써 대식세포의 항원처리 및 항원제시능을 증진시키는 것으로 보고되고 있다(Chou et al., 2020). 따라서 우리는 TLR2 및 TLR4의 작약 뿌리 추출물에 의한 p62/SQSTM1 단백질 증가에 미치는 영향을 분석하였다. 그 결과, C29에 의한 TLR2의 억제는 작약 뿌리 추출물만 처리된 그룹과 비교할 때, 작약 뿌리 추출물 매개 p62/SQSTM1의 증가에 아무런 영향이 없었으나, TAK-242에 의한 TLR4의 억제는 작약 뿌리 추출물 매개 p62/SQSTM1의 증가를 유의적으로 감소시켰다(Fig. 2A). 게다가, 작약 뿌리 추출물 매개 RAW264.7세포의 자가포식 유도도 TAK-242에 의해 TLR4가 억제된 세포에서만 감소하는 것으로 타났다. 본 결과를 미루어 볼 때, 작약 뿌리 추출물은 TLR4 의존성 p62/SQSTM1의 증가를 통해 자가포식을 유도하는 것으로 판단된다.
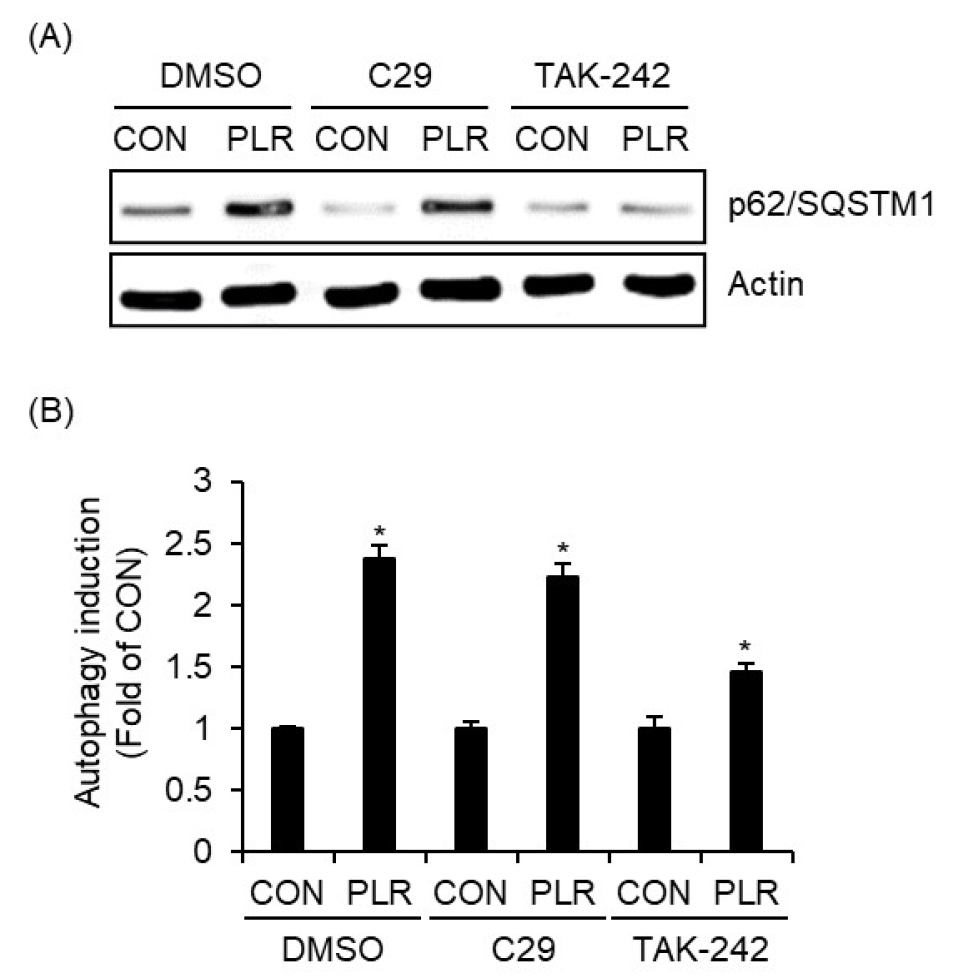
Fig. 2.
Effect of TLR2/4 on PLR-mediated increase in p62/SQSTM1 and autophagy induction. (A) RAW264.7 cells were pre-treated with C29 (100 nM) or TAK-242 (10 ㎍/mL) for 2 h and then co-treated with PLR (50 ㎍/mL) for 6 h. p62/SQSTM1 level was measured using Western blot analysis. (B) RAW264.7 cells were pre-treated with C29 (100 nM) or TAK-242 (10 ㎍/mL) for 2 h and then co-treated with PLR (50 ㎍/mL) for 24 h. Autophagy induction was measured using Autophagy Assay Kit.
작약 추출물 매개 p62/SQSTM1 단백질 증가와 관련된 신호전달 분석
작약 뿌리 추출물에 의한 p62/SQSTM1 단백질의 증가와 관련 신호전달을 구명하기 위해 RAW264.7세포에 kinase 억제제를 전처리하고 작약 뿌리 추출물을 처리한 다음 Western blot 분석으로 p62/SQSTM1 단백질의 수순을 확인하였다. 그 결과, 작약 뿌리 추출물만 처리된 그룹과 비교할 때, PD98059에 의한 ERK1/2 억제, SB203580에 의한 p38억제, SP600125에 의한 JNK 억제, 그리고 BAY 11-7082 (BAY)에 의한 NF-κB 억제는 작약 뿌리 추출물 매개 p62/SQSTM1 단백질 증가에 아무런 영향이 없었다. 그러나 LY294002에 의한 PI3K의 억제는 작약 뿌리 추출물 매개 p62/SQSTM1 단백질의 증가를 유의적으로 감소시켰다(Fig. 3A). 실제로 PI3K 신호전달은 자가포식에 의한 자가포식체 형성을 통한 병원체들의 리소솜 분해에 필수적이라고 알려져 있다(Iershov et al., 2019). 따라서, 본 결과를 미루어 볼 때, 작약 뿌리 추출물 매개 p62/SQSTM1 단백질의 증가는 PI3K의 활성화에 기인한 것으로 사료된다. 그리하여, 작약 뿌리 추출물이 PI3K를 활성화하는지 알아보기 위해, 작약 뿌리 추출물이 시간별로 처리된 RAW264.7세포에서 PI3K의 인산화를 측정하였다. 그 결과, 작약 뿌리 추출물은 처리 1시간 째부터 PI3K의 인산화를 유도하였고(Fig. 3B), 이는 작약 뿌리 추출물이 PI3K의 활성화를 유도한다는 것을 보여준다. 그리고 작약 뿌리 추출물 매개 p62/SQSTM1의 증가가 TLR4에 의존하였기 때문에, 작약 뿌리 추출물 매개 PI3K 활성화와 TLR4의 연관성을 조사하기 위해, TAK-242로 TLR4를 억제시킨 후 작약 뿌리 추출물을 처리하고 PI3K의 활성화 여부를 조사하였다. 그 결과, 작약 뿌리 추출물만 처리된 그룹에 비해 TAK-242에 의한 TLR4의 억제는 작약 뿌리 추출물 매개 PI3K의 인산화를 감소시켰다(Fig. 3C). 본 결과들을 미루어 볼 때, 작약 뿌리 추출물은 TLR4/PI3K의 활성화를 통해 p62/SQSTM1의 증가를 유도하는 것으로 사료된다.

Fig. 3.
Effect of kinases on PLR-mediated increase in p62/SQSTM1. (A) RAW264.7 cells were pre-treated with PD98059 (40 μM), SB203580 (40 μM), SP600125 (40 μM), BAY 11-7082 (BAY, 20 μM) or LY294002 (40 μM) for 2 h and then co-treated with PLR (50 ㎍/mL) for 6 h. p62/SQSTM1 level was measured using Western blot analysis. (B) RAW264.7 cells were treated with PLR (50 ㎍/mL) for the indicated times. p-PI3K and PI3K levels were measured using Western blot analysis. (C) RAW264.7 cells were pre-treated with TAK-242 (10 ㎍/mL) for 2 h and then co-treated with PLR (50 ㎍/mL) for 1 h. p-PI3K level was measured using Western blot analysis.
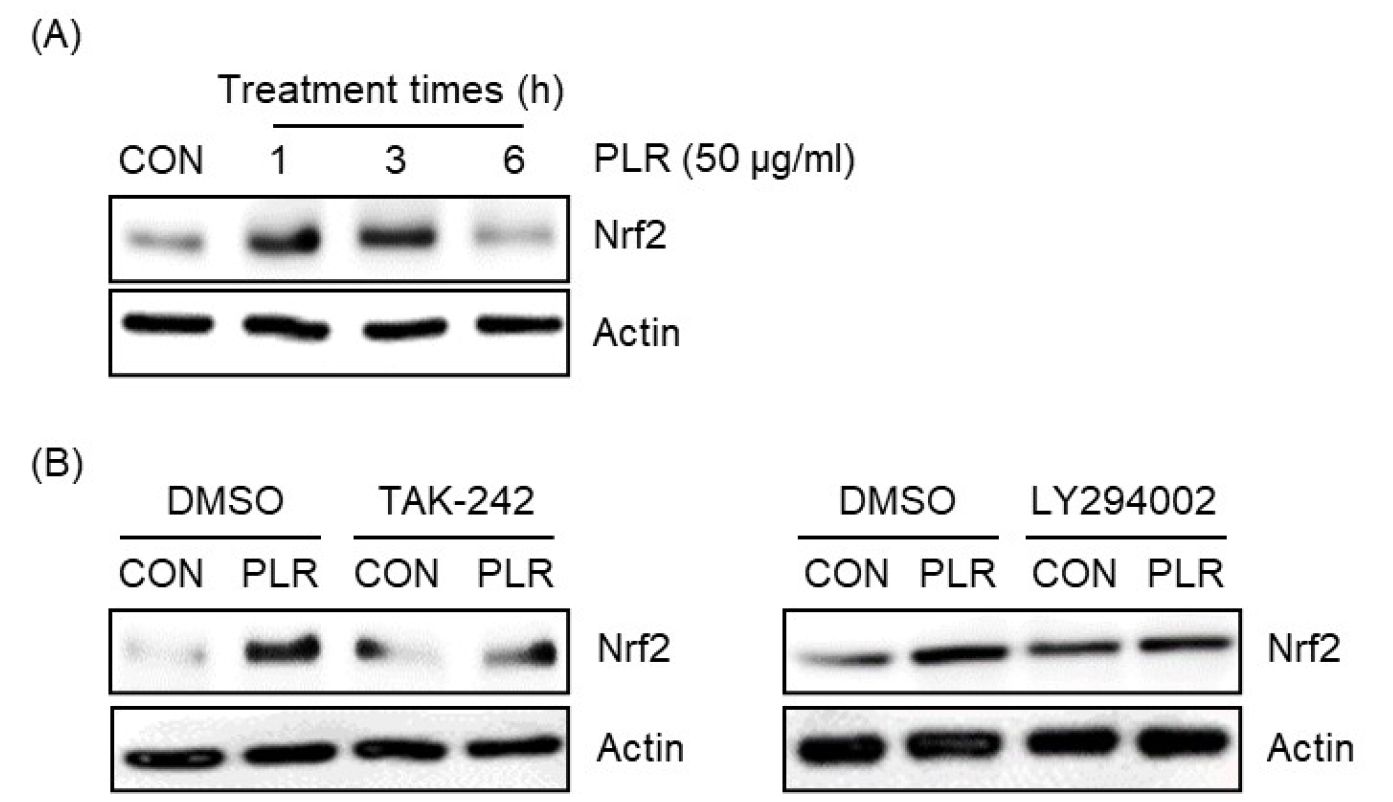
Nrf2가 작약 뿌리 추출물 매개 p62/SQSTM1 단백질의 증가에 미치는 영향
Nrf2는 p62/SQSTM1의 발현을 증가시켜 자가포식 과정에 관여함으로 선천성 면역반응의 활성화에 관여한다고 보고되었다(Fujita et al., 2011a, 2011b; Thimmulappa et al., 2006). 그리하여 작약 뿌리 추출물이 Nrf2의 발현에 미치는 영향을 조사한 결과, 작약 뿌리 추출물이 처리된 RAW264.7 세포에서 Nrf2가 증가하는 것으로 나타났다(Fig. 4A). 작약 뿌리 추출물 매개 p62/SQSTM1의 증가가 TLR4/PI3K 신호전달 활성화에 의존하였기 때문에, TLR4와 PI3K가 작약 뿌리 추출물 매개 Nrf2 증가에 미치는 영향을 조사하였다. 그 결과, TAK-242에 의한 TLR4 억제와 LY294002에 의한 PI3K의 억제는 작약 뿌리 추출물 매개 Nrf2의 증가를 유의적으로 감소시켰다(Fig. 4B). 따라서 본 결과들은 작약 뿌리 추출물은 TLR4/PI3K의 신호전달 활성화를 통해 Nrf2를 증가시키는 것으로 사료된다.