

Introduction
Materials and Methods
Experimental materials and growth conditions
Leaf growth characteristics investigation
Physiological measurements
Data analysis
Results
Growth environment
Differences in leaf mass per area (LMA) under shading treatments
Differences in chlorophyll content under shading treatments
Differences in chlorophyll fluorescence response under shading treatments
Differences in photosynthetic characteristics under shading treatments
Differences in stomatal conductance, transpiration rate, and water use efficiency under shading treatments
Discussion
Shading treatment and growth environment
Photomorphogenic adaptation responses of D. maackianum to shading treatments
Physiological adaptation responses of D. maackianum to shading treatment
Analysis of correlations between light intensity and growth and physiological traits
Introduction
Delphinium maackianum Regel, a perennial herb in the Ranunculaceae family, is distributed across the Korean Peninsula, China, and Russia. In Korea, it is found along the Baekdudaegan mountain range in Gangwon, Gyeonggi, Gyeongsangbuk, and Chungcheongbuk provinces. With a typical height of about 1 meter, it blooms dark purple flowers in racemes from July to September (Lee, 2014). This species thrives in rich, moist forest soils and is known to be sensitive to high temperatures and direct sunlight (KNA, 2024). Historically, Delphinium species have been used in traditional medicine to treat gastrointestinal ailments and seizures, and they are recognized for their anti-inflammatory, anticancer, and cosmetic benefits (Chung, 2016; Jung et al., 2006; Kim et al., 2023; Kwon et al., 2009). Additionally, the striking blue flowers of D. maackianum make it a popular ornamental plant in Korean gardens (Sohn, 2013).
Despite its aesthetic and medicinal value, D. maackianum is classified as Near Threatened (NT) on the national red list due to its limited Extent of Occurrence (EOO) and Area of Occupancy (AOO). This plant is highly valued not only for its striking dark purple flowers, which make it a popular choice in ornamental gardening, but also for its traditional medicinal uses in treating gastrointestinal ailments and seizures. Additionally, it possesses recognized anti-inflammatory, anticancer, and cosmetic properties. However, its high value has led to extensive exploitation, resulting in significant pressure on natural populations.
Efforts to conserve D. maackianum have often been hindered by a lack of detailed physiological data necessary to create suitable conservation conditions. The species is particularly vulnerable to threats from habitat disturbance and over-collection, especially as many of its habitats are located near forest roads, which exposes them to human interference and trampling (KNA, 2021). As a boreal plant, its southern distribution limit lies within Korea, making it especially susceptible to rapid environmental changes, such as those induced by climate change (Gantsetseg et al., 2020). These factors put D. maackianum at significant risk, and current conservation measures have proven insufficient to ensure its survival. This situation underscores the urgent need for both in situ and ex situ conservation strategies that are informed by comprehensive physiological data.
Given these challenges, research focused on understanding the physiological and ecological requirements of D. maackianum is critical. Such research is essential for developing optimal growth conditions that can support effective ex situ conservation. Without this foundational data, conservation efforts will continue to fall short, leaving this valuable species at risk of further decline.
Botanical gardens play a crucial role in plant diversity conservation. Their exhibition gardens, with various microclimates, can serve as ideal sites for the ex situ conservation of multiple plant species (Price et al., 2004; Smith, 2019; Thomas et al., 2022). Effective ex situ conservation requires planting target species in areas within these gardens that offer optimal microclimatic conditions. Understanding the physiological and ecological responses of plants to environmental factors such as light, temperature, and moisture is essential for determining these optimal conditions (Larcher, 2003). Light conditions, in particular, are critical as they are a primary energy source and regulator of plant growth and development. Plants have evolved to optimize photosynthesis based on their light environment, but growth and physiological activities can be hindered if light conditions exceed their adaptive capacity (Folta and Carvalho, 2015; Kim et al., 2013; Paradiso and Proietti, 2022). Thus, investigating the optimal light environment for plant growth is vital for successful ex situ conservation.
Currently, there is a lack of research on the ex situ conservation of D. maackianum, and no studies have been conducted to determine the optimal light conditions for its growth. Therefore, this study aims to investigate and analyze the leaf growth and physiological responses of D. maackianum under various light conditions to identify the optimal light environment for its growth, contributing to effective ex situ conservation.
Materials and Methods
Experimental materials and growth conditions
The plant materials used in this experiment were seedlings that had grown for two months’ post-germination. Seeds stored at 4°C in the National DMZ Botanical Garden were germinated in a greenhouse in May 2023. After one month of growth, the seedlings were transplanted into 15 ㎝ diameter pots (Square Large Pot; Hawbunplaza Co., Ltd., Hanam, Korea) filled with a soil mixed with akadama soil and horticultural soil (Hanareum No.2; Shinsung mineral Co., Ltd., Goesan, Korea) at 1 : 2 (v : v). These plants were grown in the greenhouse until early July 2023 before being moved to the experimental plots. The experimental plots were located within the National DMZ Botanical Garden and consisted of three plots each with dimensions of W2,000 × D14,000 × H2,000. To ensure the soil in the pots remained moist, bottom watering was employed. For shading treatments, double- layered vinyl shade nets (UV Large Shade net; Hansol Co., Ltd., Boseong, Korea) were used to create shading levels of 0% (control), 50%, 75%, and 95%. Each treatment group included six replicates, resulting in a total of 24 plants for the shading experiment. To ensure ventilation, the bottom of the shade nets was raised 20 ㎝ above the ground. From July to September 2023, the plants were grown under different light conditions, and their growth and physiological differences were investigated.
To compare the differences in growth conditions under different shading treatments, temperature and humidity data loggers (HOBO U23 Pro v2 U23-001; Onset; Boume, MA, USA) were installed inside both the control and shaded plots to monitor air temperature and humidity. Light intensity was measured using a quantum sensor (Quantum Sensor, Apogee, USA) connected to a data logger (MSTL Mini Logger, STA Corp., Korea), and light quality was measured using a portable spectroradiometer (LI-180, Li-Cor Inc., USA).
Leaf growth characteristics investigation
To investigate the leaf growth characteristics of D. maackianum under different shading levels, six healthy leaves were collected from each treatment group to measure the Leaf Mass per Area (LMA). Leaf area was measured using a leaf area meter (LI-3100C, Li-Cor Inc., USA). The collected leaves were then dried in a forced-air dryer at 80°C for 48 hours before their dry weight was measured using an electronic balance. The LMA was calculated using the following formula:
Leaf Mass per Area (LMA) = Leaf Mass/Leaf Area (㎎·㎝-2)
Physiological measurements
Leaf chlorophyll content
To measure chlorophyll content, three leaves were collected from each treatment group. Leaf discs, each with an area of 0.72 ㎠, were cut from the leaves and placed in vials containing 5 ml of DMSO (Dimethyl sulfoxide). The vials were then incubated at 65°C for 6 hours to extract the pigments (Hiscox and Israelstam, 1979). The absorbance of the extracted solution was measured at 665.1 nm and 649.1 nm using a spectrophotometer (UV/VIS Spectrophotometer- 2550, Shimadzu Inc., Japan). The chlorophyll a and b contents were calculated using the formulas from Wellburn (1994):
Chlorophyll a = 12.47 A665.1 - 3.62 A649.1
Chlorophyll b = 25.06 A649.1 - 6.50 A665.1
Chlorophyll fluorescence response
Chlorophyll fluorescence responses, including the maximum quantum yield (Fv/Fm) and non-photochemical quenching (NPQ), were measured using a chlorophyll fluorometer (PAM-2500, Heinz Walz GmbH, Germany). Five plants showing average growth were selected from each treatment group. Before measurement, the leaves were dark-adapted for about 20 minutes using sample clips to deactivate the photosynthetic heat dissipation pathways. Fluorescence intensity was measured using the quenching analysis method, which involved sequentially illuminating the leaves with measuring light and saturating pulses for 200 seconds. Fo was measured using a light intensity below 0.1 μmol·m-2·s-1, while Fm was measured using a saturating light pulse of 8,000 μmol·m-2·s-1 for 0.8 seconds. The photochemical reactions were calculated using the formulas from Lichtenthaler et al. (2005).
Photosynthetic characteristics
To investigate photosynthetic characteristics, five plants showing average growth were selected from each treatment group. To prevent photosynthetic variations due to water stress, the plants were watered a day before measurement. Additionally, they were dark-adapted using black vinyl covers to avoid errors due to light exposure. A portable photosynthesis system (LI-6400XT, Li-Cor Inc., USA) was used to generate light-response curves and CO2-response curves, following the method of Kim and Lee (2001).
For the light-response curves, the Photosynthetic Photon Flux Density (PPFD) was adjusted to 0, 25, 50, 75, 100, 150, 300, 500, 700, 1000, and 1500 μmol·m-2·s-1. The incoming CO2 concentration was set at 400 μmol·(CO2)·m-2·s-1 air, and the chamber temperature was maintained at 20°C. From the light-response curves, parameters such as dark respiration rate (Rd), apparent quantum yield (AQY), light compensation point (LCP), light saturation point (LSP), and maximum photosynthetic rate (PNmax) were calculated.
For the CO2-response curves, the ambient CO2 concentration was adjusted to 0, 50, 100, 200, 300, and 400 μmol·(CO2)·m-2·s-1 air, with PAR maintained at 1,000 μmol·m-2·s-1. Photosynthesis and intercellular CO2 concentration (Ci) were measured to calculate parameters such as photorespiration rate (Rp), CO2 compensation point (CCP), and carboxylation efficiency (CE).
Stomatal conductance and transpiration rate were calculated using the formulas from von Caemmerer and Farquhar (1981), and water-use efficiency was calculated using the formula from Kim et al. (2002).
WUE = Photosynthetic rate/Transpiration rate
Data analysis
In this experiment, the statistical analysis of the results from different shading treatments was conducted using Scheffe’s significant difference test to determine the significance of the differences between groups. Post-hoc comparisons were performed using Dunnett’s T3 test, with a significance level set at p=0.05. Additionally, correlation analyses were conducted to examine the relationships between the attributes of plants within each shading treatment group. To assess the normality of the data within each group, the Shapiro-Wilk test was employed. All statistical analyses were carried out using SPSS version 21.0 (IBM Inc., USA).
Results
Growth environment
To observe the physiological characteristics of D. maackianum under different shading treatments, experimental plots were set up with full light conditions (control) and shading treatments at 50%, 75%, and 95%. Using temperature and humidity data loggers (HOBO U23 Pro v2 U23-001; Onset; Bourne, MA, USA), the ambient temperature and humidity were measured for each treatment group, as shown in Table 1. During the experimental period, the average day and night temperatures were highest in the control group at 21.63°C and 17.32°C, respectively. The lowest temperatures were observed in the 75% shading treatment at 20.27°C during the day and 16.61°C at night. The maximum daily temperature was highest in the control group at 34.55°C, while the minimum daily temperature was lowest in the 75% shading treatment at 2.61°C. Relative humidity was highest in the 95% shading treatment at 91.44% and lowest in the control group at 86.51%. To compare the actual light intensity differences due to shading installations, a Quantum Sensor (Apogee, USA) was used to measure the light levels (Table 2). The relative light intensity compared to full light conditions was 50.9% for the 50% shading treatment, 26.2% for the 75% shading treatment, and 12.0% for the 95% shading treatment. Notably, the actual light level in the 95% shading treatment was 7% higher than the marketed specification. A portable spectroradiometer (LI-180, Li-Cor Inc., USA) was used to measure the light quality, specifically the red to far-red light ratio (R/FR), within each shading treatment (Table 2). Both the control and the 50% shading treatment had an R/FR ratio of 1.1. In the 75% and 95% shading treatments, the R/FR ratio was 1.0, showing no significant differences in the light quality across these shading levels.
Table 1.
Temperature and relative humidity following each shaded treatment.
Table 2.
Relative light intensity (RLI), light quality, and resulting leaf mass per area (LMA) of Delphinium maackianum Regel in sunlight and each shaded treatment plot. R/FR is the ratio of infrared to far infrared.
| Light condition | Sunlight | Shaded 50% | Shaded 75% | Shaded 95% | p |
| RLI (%) | 100.0 | 50.9 | 26.2 | 12.0 | |
| R/FR | 1.1 | 1.1 | 1.0 | 1.0 | n.s. |
| LMA (㎎/㎠) | - | 9.3±1.6az | 9.9±1.0a | 8.5±0.6a | * |
Differences in leaf mass per area (LMA) under shading treatments
The ratio of infrared to far-infrared radiation remained consistent between 1.0 and 1.1 across all treatments. The Leaf Mass per Area (LMA) of D. maackianum was measured under different shading levels (Table 2). Leaf Mass Area (LMA) data for D. maackianum varied across different shading treatments. LMA was not measurable under 100% light condition due to plant mortality. The LMA was significantly lower in the 95% shading treatment group, with a value of 8.5 ± 0.6 ㎎·㎠, compared to other treatments. The highest LMA was observed in the 75% shading treatment group, with a value of 9.9 ± 0.1 ㎎·㎠. Statistical analysis revealed no significant differences in LMA among the shading treatments (p > 0.05). However, there were slight differences observed, with LMA showing a trend of decreasing from 50% shading to 95% shading conditions.
Differences in chlorophyll content under shading treatments
To assess the effect of shading on chlorophyll content in D. maackianum, the chlorophyll a, b, and a+b contents were measured for each treatment group (Fig. 1). The 95% shading treatment group had significantly higher chlorophyll a, b, and total chlorophyll (a+b) contents compared to the other groups. Although there was a trend of increasing chlorophyll content with higher shading levels, there was no significant difference between the 50% and 75% shading treatment groups. The chlorophyll a/b ratio did not show significant differences across shading levels.
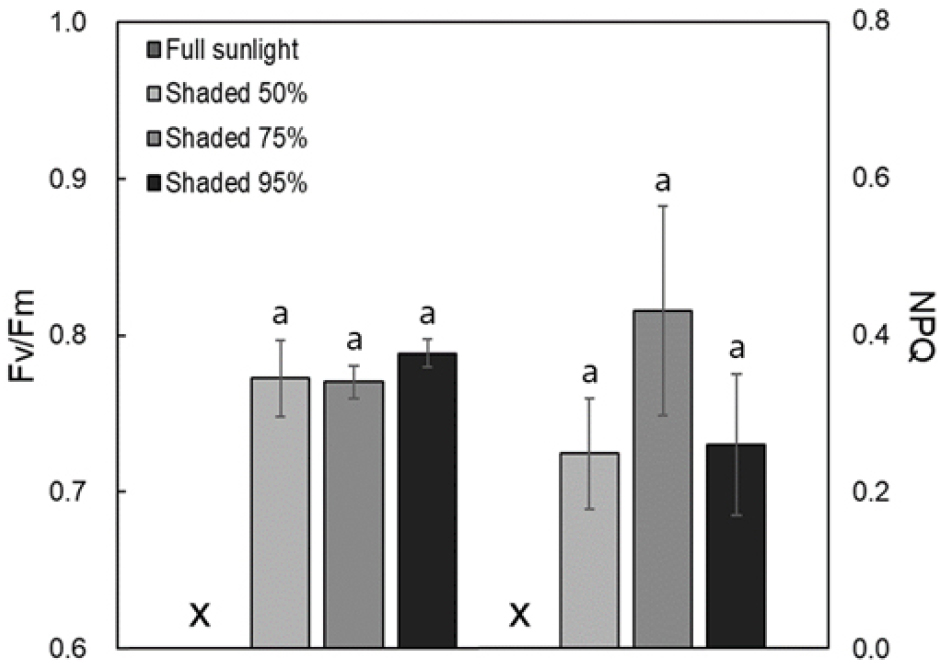
Fig. 1.
Chlorophyll contents in Delphinium maackianum Regel under different shading treatments. Chlorophyll a, b, and a+b contents were highest in the 95% shading treatment. Chlorophyll a/b did not differ among shading treatments. Statistical significance was set at p < 0.05 and vertical bars represent standard deviations (n=3).
Differences in chlorophyll fluorescence response under shading treatments
The effects of shading on chlorophyll fluorescence parameters such as maximum quantum yield (Fv/Fm) and non-photochemical quenching (NPQ) were measured (Fig. 2). The Fv/Fm value was highest in the 95% shading treatment group at 0.79 ± 0.01, but the differences among treatment groups were not statistically significant. The NPQ was highest in the 75% shading treatment group, yet again, no significant differences were found among the treatments.
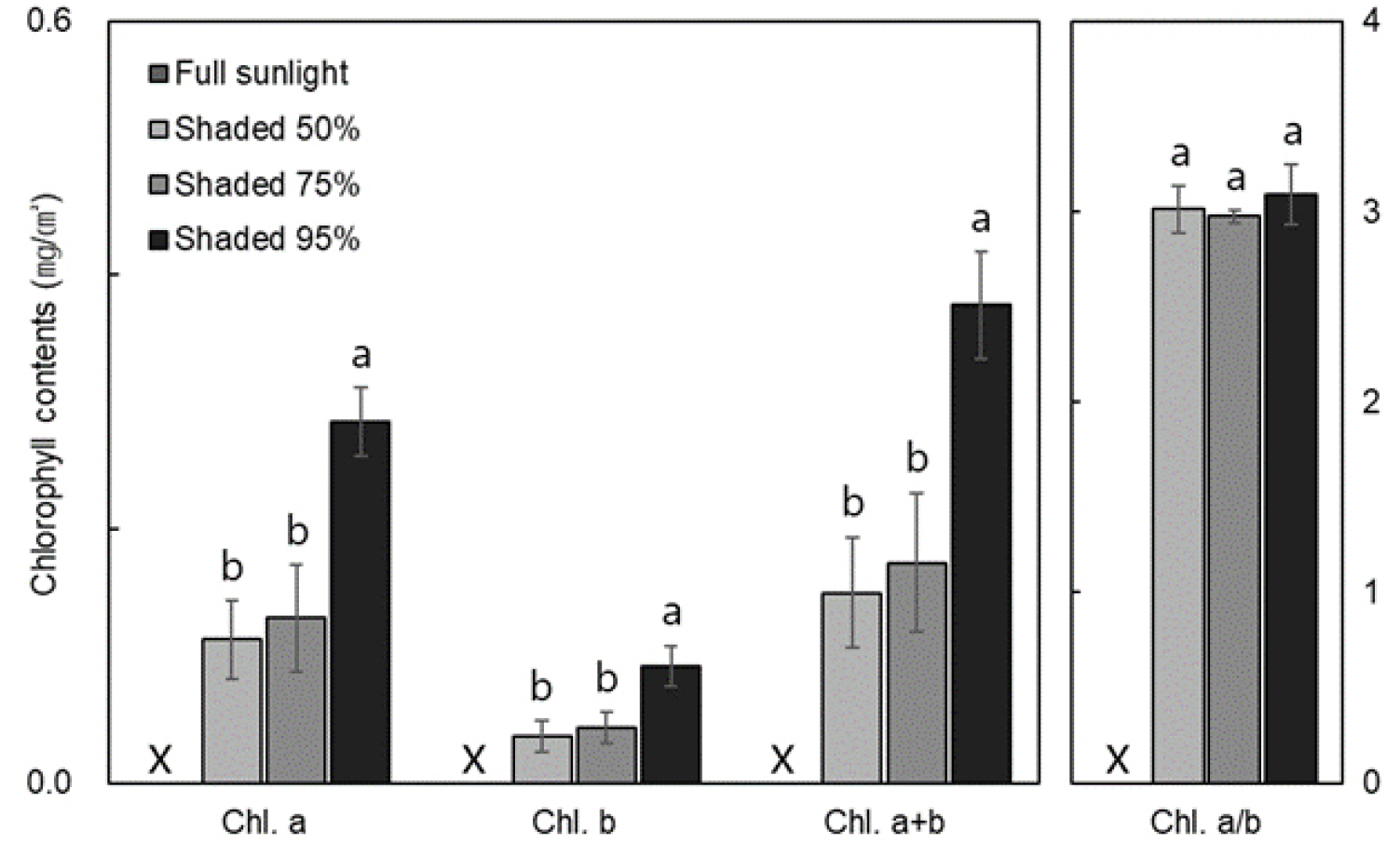
Fig. 2.
Maximum quantum efficiency of PSII photochemistry (Fv/Fm) and Nonphotochemical chlorophyll fluorescence quenching (NPQ) of Delphinium maackianum Regel under different shading treatments. Both Fv/Fm and NPQ were not significantly different shading treatments. Statistical significance was set at p < 0.05 and vertical bars represent standard deviations (n=6).
Differences in photosynthetic characteristics under shading treatments
Photosynthetic responses of D. maackianum under different shading levels were evaluated by constructing light response curves and CO2 response curves. Various photosynthetic parameters such as dark respiration rate (Rd), apparent quantum yield (AQY), light compensation point (LCP), light saturation point (LSP), and maximum photosynthetic rate (PNmax) were assessed (Fig. 3a and Table 3).
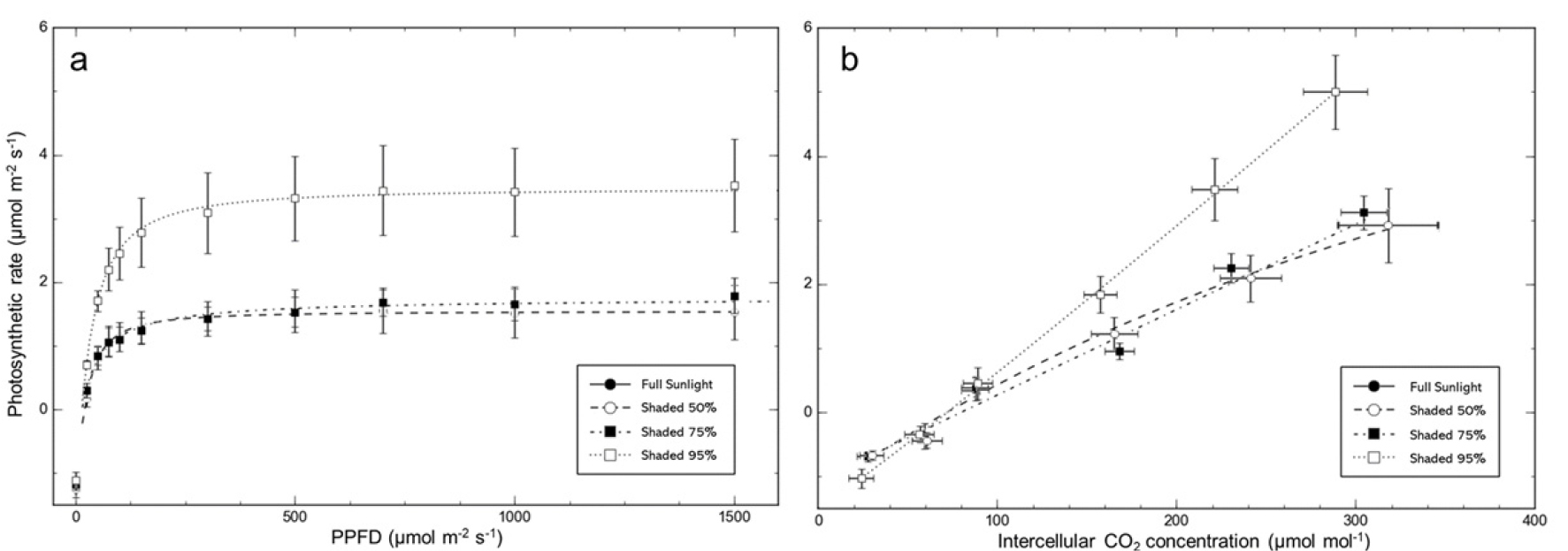
Fig. 3.
Photosynthesis curves (a), and A-Ci curves (b) of Delphinium maackianum Regel under different shading treatments. The photosynthetic rate was highest at 95% shading, but lowest at 50% shading. The photosynthetic rate of D. maackianum in response to CO2 concentration was higher in 95% shading compared to 50% and 75% shading. Vertical bars represent standard deviations (n=5).
Table 3.
Photosynthetic characteristics of Delphinium maackianum Regel in response to shading treatments estimated from photosynthesis curves. Light compensation point was lowest at 95% shading, while light saturation point and photosynthetic rate, apparent quantum yield were highest at 95% shading.
| Itemz | Shade Level (%) | |||
| 0 | 50 | 75 | 95 | |
| Rd | - | 1.25ay | 1.18a | 1.11a |
| (±0.13) | (±0.08) | (±0.13) | ||
| LCP | - | 17.25a | 13.20ab | 10.80b |
| (±2.23) | (±2.40) | (±1.47) | ||
| LSP | - | 181.50b | 237.60ab | 307.20a |
| (±36.32) | (±43.24) | (±51.77) | ||
| PNmax | - | 1.58b | 1.80b | 3.53a |
| (±0.37) | (±0.27) | (±0.73) | ||
| AQY | - | 0.02b | 0.02b | 0.04a |
| (±0.00) | (±0.00) | (±0.01) | ||
zDark respiration; Rd [μmol (CO2) m-2 s-1], light compensation point; LCP [μmol m-2 s-1], light saturation point; LSP [μmol m-2 s-1], maximum photosynthesis rate; PNmax [μmol (CO2) m-2 s-1], net apparent quantum yield; AQY [mmol (CO2) mol-1]. The assumption of homogeneity of variance was assessed with Levene’s test.
The 95% shading treatment exhibited the lowest dark respiration rate (Rd) at 1.11 ± 0.13 μmol·(CO2)·m2·s-1, though the differences among treatments were not statistically significant. The light compensation point (LCP) was significantly higher in the 50% shading treatment at 17.25 ± 2.23 μmol·m2·s-1 and decreased as shading increased. Conversely, the light saturation point (LSP) peaked at 307.20 ± 51.77 μmol·m2·s-1 in the 95% shading treatment, with lower shading levels showing a decline. Both the maximum photosynthetic rate (PNmax) and apparent quantum yield (AQY) were significantly elevated in the 95% shading treatment, decreasing with reduced shading levels, though no significant differences were observed between the 50% and 75% shading treatments.
To further understand the impact of shading on photosynthetic responses to internal leaf CO2 concentration (Ci), CO2 response curves were constructed (Fig. 3b and Table 4). Parameters such as photorespiration rate (Rp), CO2 compensation point (CCP), and carboxylation efficiency (CE) were derived.
Table 4.
Photosynthetic characteristics of Delphinium maackianum Regel in response to shading treatments estimated from A-Ci curves. Photo-respiration rate and carboxylation efficiency were highest at 95% shading. Meanwhile, the CO2 compensation point was not significantly different between shading treatments.
| Itemz | Shade Level (%) | |||
| 0 | 50 | 75 | 95 | |
| Rp | - | 1.12by | 0.94b | 1.54a |
| (±0.19) | (±0.12) | (±0.14) | ||
| CCP | - | 71.63a | 75.49a | 71.20a |
| (±5.46) | (±6.31) | (±6.47) | ||
| CE | - | 0.01b | 0.01b | 0.02a |
| (±0.00) | (±0.00) | (±0.00) | ||
The highest photorespiration rate (Rp) was observed in the 95% shading treatment at 1.12 ± 0.19 μmol (CO2)·m2·s-1, with no significant differences between the 50% and 75% shading treatments. Similarly, carboxylation efficiency (CE) was highest in the 95% shading treatment at 0.02 ± 0.00 μmol·(CO2)·mol-1, with no significant differences between the 50% and 75% treatments. There were no significant differences in the CO2 compensation point (CCP) among the treatment groups.
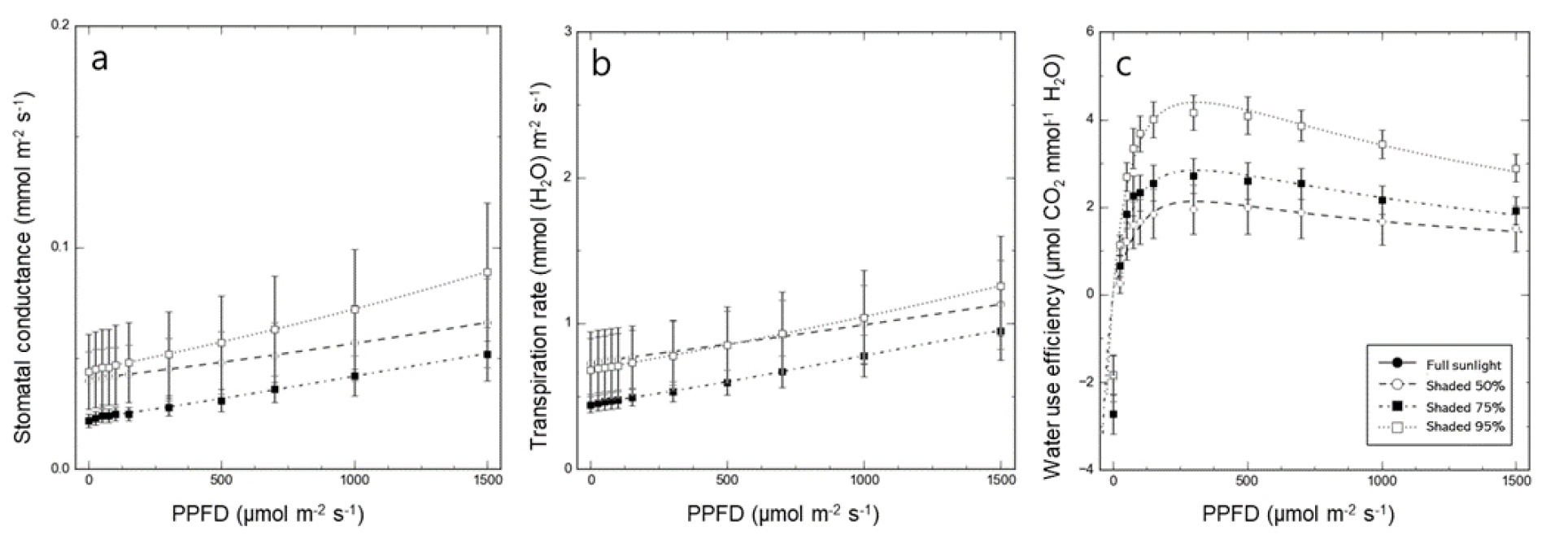
Differences in stomatal conductance, transpiration rate, and water use efficiency under shading treatments
transpiration rate, and water use efficiency (WUE) were also investigated (Fig. 4). Both stomatal conductance and transpiration rate were lowest in the 75% shading treatment and highest in the 95% shading treatment. The WUE was highest in the 95% shading treatment and lowest in the 50% shading treatment.
Discussion
Shading treatment and growth environment
As the shading level increased, the ambient temperature tended to decrease while the relative humidity increased. This is likely because higher shading levels reduce direct sunlight exposure, resulting in lower temperature increases and reduced transpiration, leading to higher relative humidity.
In forest stands, light that reaches the understory through the canopy has a lower intensity compared to full sunlight, and the red light wavelength is absorbed in the upper layers, altering the R/FR ratio, which affects plant growth (Skálová et al., 1999). In this experiment, there were no significant differences in light quality among the treatments, suggesting that light quality did not affect the results of the experiment. In the control group under full sunlight, all plants died. The excessive light and high temperatures likely caused excessive transpiration, leading to water imbalance and leaf scorch, which ultimately resulted in plant death (Je et al., 2006).
Photomorphogenic adaptation responses of D. maackianum to shading treatments
Photomorphogenic adaptation refers to the changes in the distribution of substances within a plant’s organs and structures to optimize photosynthesis under given light conditions (Folta and Carvalho, 2015; Kim et al., 2008; Paradiso and Proietti, 2022). The morphological characteristics of leaves, which are key photosynthetic organs, are closely related to processes such as photosynthesis and respiration (Chim and Han, 2003). Leaf Mass per Area (LMA) is a measure of the weight of a leaf per unit area and explains the relationship between light absorption, growth, and carbon acquisition in plants (Cheng et al., 2014).
From a photomorphogenic perspective, plants generally exhibit thinner leaves with larger surface areas and lower LMA under low light conditions to maximize light capture (Salisbury and Ross, 1992). In this study, D. maackianum showed the lowest LMA in the 95% shading treatment. However, there were no significant differences among the treatments, indicating that the plants did not exhibit photomorphogenic adaptation responses to different light intensities. Despite the lack of statistical significance, this trend suggests a potential adaptation of D. maackianum to varying light intensities by adjusting its leaf mass per unit area. The consistent ratio of infrared to far-infrared radiation across all treatments indicates that differences in shading did not significantly alter the spectral composition of the light reaching the plants. This suggests that the observed variations in LMA are more likely influenced by the intensity rather than the spectral quality of light. While statistical significance was not reached, the slight decrease in LMA with increased shading levels hints at a nuanced response of D. maackianum to light availability (Jo et al., 2024; Yun et al., 2024). Further studies could explore additional physiological and biochemical parameters to elucidate the adaptive strategies of this species under varying light conditions.
Physiological adaptation responses of D. maackianum to shading treatment
Plants optimize photosynthesis by adjusting physiological characteristics such as chlorophyll content, chlorophyll fluorescence response, and photosynthetic traits based on their light environment. This is referred to as physiological adaptation responses (Kim et al., 2008; Paradiso and Proietti, 2022). Total chlorophyll content is a physiological indicator of photosynthetic capacity and is influenced by light conditions, affecting the plant’s shade tolerance (Kim et al., 2001; Mauro et al., 2011; Verhoeven et al., 2005). Shade- tolerant species typically have higher chlorophyll content under low light to increase light use efficiency and carbon acquisition (Valladares and Niinemets, 2008). Conversely, under high light, shade-tolerant species reduce chlorophyll content to prevent photoinhibition by decreasing light absorption through Photosystem II (Je et al., 2006; Kyparissis et al., 2000).
In this experiment, the chlorophyll a, b, and total chlorophyll content increased with higher shading levels, indicating that D. maackianum exhibits typical shade plant responses by increasing chlorophyll content under low light to enhance light use efficiency and decreasing it under high light for photoprotection. Chlorophyll a/b ratio is used as an indicator of leaf aging and shade adaptation, typically decreasing in high shade-tolerant plants under low light environments due to increased chlorophyll b for stabilizing light-harvesting complex proteins (Fails et al., 1982; Kim et al., 2003; Kim et al., 2020; Lee et al., 2019). However, no significant differences were observed in the chlorophyll a/b ratio among the shading treatments, likely due to the lack of variation in light quality across treatments.
Photosynthesis is sensitive to environmental stress (Maxwell and Johnson, 2000). When chlorophyll absorbs excess photons, it initiates energy dissipation pathways to relieve excited states, with heat and fluorescence reactions serving as key indicators of plant stress (Maxwell and Johnson, 2000; Rascher et al., 2000). Parameters such as maximum quantum yield (Fv/Fm) and non-photochemical quenching (NPQ) are commonly used to assess stress (Baker, 2008; Strauss et al., 2006). Fv/Fm represents the maximum quantum yield of Photosystem II in dark-adapted plants and indicates photosystem activity and photosynthetic capacity, typically decreasing with increased stress (Baker, 2008; Borkowska, 2002; Lawson et al., 2008). NPQ measures the dissipation of excess light energy as heat, usually increasing under stress conditions (Genty et al., 1990). For D. maackianum, Fv/Fm was highest in the 95% shading treatment and NPQ in the 75% treatment, but neither showed significant differences among treatments, suggesting that chlorophyll content reduction under high light prevented photoinhibition, maintaining the functionality of the photosynthetic apparatus. However, further considerations on the photosynthetic fluorescence response are expected to be necessary in the future.
Dark respiration rate (Rd) indicates the energy acquisition through respiration essential for plant survival and growth (Noguchi et al., 1996). Shade-tolerant plants generally exhibit lower Rd and light compensation points under low light to efficiently utilize light and maintain high photosynthetic rates (Lee et al., 2013; Walters et al., 1993). In this study, D. maackianum showed a significant decrease in light compensation point with increased shading, aligning with studies suggesting that shade-tolerant plants have light compensation points of 5-10 μmol·m-2·s-1 (Larcher, 2003). The significantly higher light saturation point in the 95% shading treatment further indicates enhanced photosynthesis under low light, demonstrating effective physiological adaptation responses.
Apparent quantum yield (AQY) measures the efficiency of converting light energy into photochemical energy under low light, serving as an indicator of photosynthetic capacity (Evans, 1987; Kim and Lee, 2001). D. maackianum had the highest AQY in the 95% shading treatment, mirroring the trend in maximum photosynthetic rate, suggesting increased activation of the photochemical system under low light. The high photorespiration rate and carboxylation efficiency in the 95% shading treatment also indicate enhanced carbon fixation capacity, demonstrating physiological adaptation responses.
Plants regulate stomatal conductance to control gas exchange with the atmosphere, influencing transpiration rate, photosynthesis, and water use efficiency (Hinckley and Braatne, 1994; Lee et al., 2012; Lim et al., 2006). In this study, D. maackianum exhibited higher stomatal conductance and transpiration rates under 95% shading, leading to active gas exchange and high water use efficiency, promoting photosynthesis. Conversely, higher light levels in the 50% and 75% shading treatments reduced stomatal conductance and transpiration rates, especially under 50% shading, resulting in significantly lower water use efficiency (Farooq et al., 2009; Lee et al., 2013).
Analysis of correlations between light intensity and growth and physiological traits
To analyze the correlations between relative light intensity, leaf growth characteristics, and physiological traits under different shading levels, a Pearson correlation analysis was performed (Fig. 5). The results showed negative correlations between relative light intensity and chlorophyll a, b, total chlorophyll content, light compensation point, light saturation point, maximum photosynthetic rate (p<0.001), apparent quantum yield (p<0.01), CO2 compensation point, and photorespiration rate (p<0.05). This indicates that under lower light conditions, D. maackianum exhibits physiological adaptation responses to optimize photosynthesis by increasing light absorption and utilization efficiency (Paradiso and Proietti, 2022).
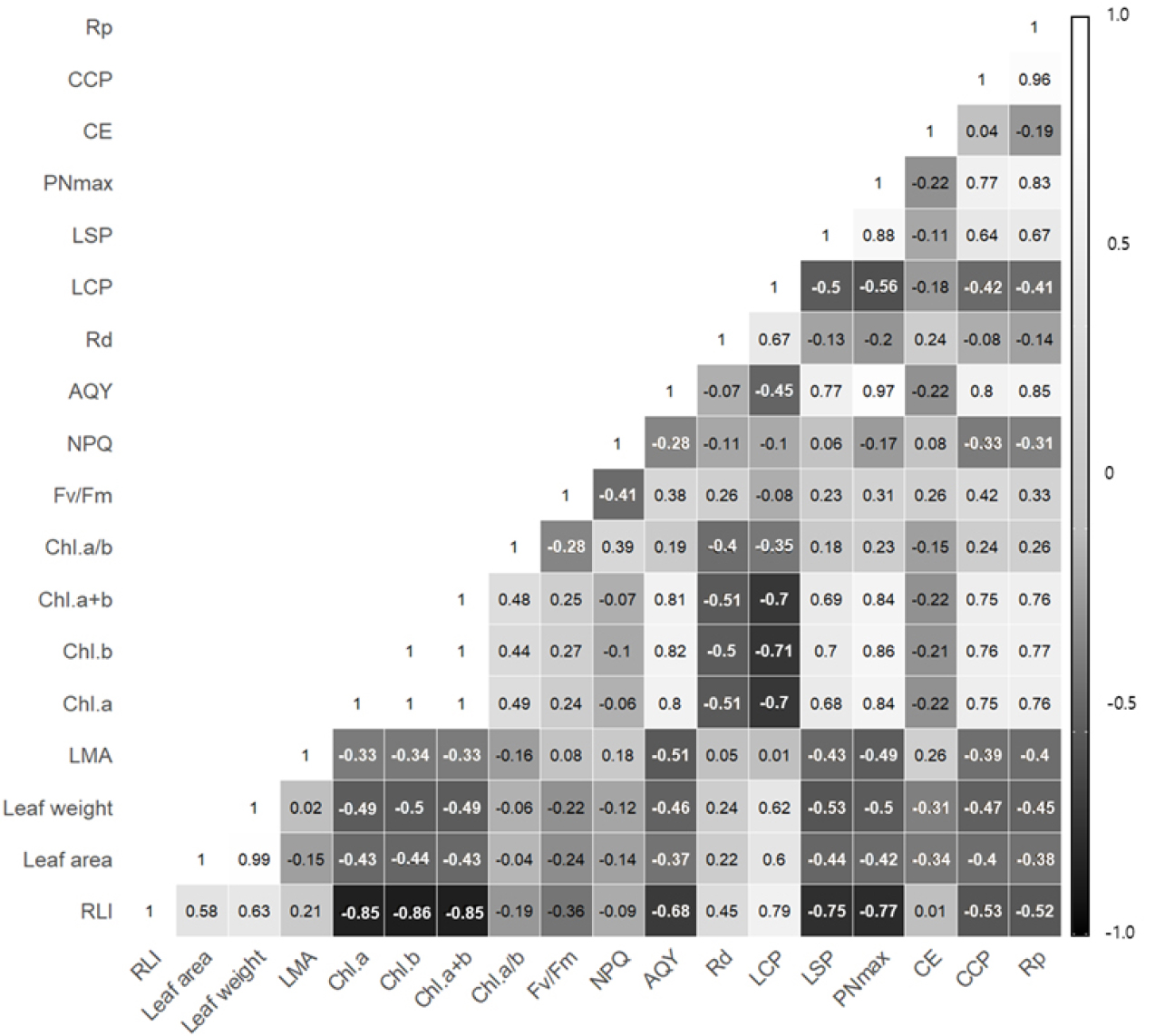
Fig. 5.
Heatmap of Pearson correlation analysis of growth and physiological characteristics of Delphinium maackianum Regel under different shading treatments. Closer to white indicates a positive correlation and closer to black indicates a negative correlation. (Relative light intensity; RLI, leaf mass per area; LMA, chlorophyll contents; Chl, maximum quantum yield; Fv/Fm, non-photochemical quenching; NPQ, net apparent quantum yield; AQY, dark respiration; Rd, light compensation point; LCP, light saturation point; LSP, maximum photosynthesis rate; PNmax, photo-respiration rate; Rp, CO2 compensation point; CCP, carboxylation efficiency; CE).
Furthermore, total chlorophyll content was positively correlated with apparent quantum yield, maximum photosynthetic rate, CO2 compensation point, photorespiration rate (p<0.001), and light saturation point (p<0.01). This suggests that higher chlorophyll content is associated with increased photosynthetic activity. The maximum photosynthetic rate showed a negative correlation with the light compensation point (p<0.05) and positive correlations with apparent quantum yield and light saturation point (p<0.001). These findings align with the response of plants to maximize net photosynthesis by lowering the light compensation point and increasing the light saturation point and maximum photosynthetic rate under given light conditions (Lee et al., 2013; Walters et al., 1993).
In the context of the carbon fixation system, carboxylation efficiency did not show significant correlations with other traits, but the CO2 compensation point and photorespiration rate were negatively correlated with relative light intensity (p<0.05) and positively correlated with chlorophyll content, apparent quantum yield, maximum photosynthetic rate (p<0.001), and light saturation point (p<0.01). Although carboxylation efficiency is an indicator of rubisco enzyme activity and content, the study results did not match the findings that higher carboxylation efficiency enhances photosynthetic function (Reddy et al., 2004). Instead, the trend observed was that a lower CO2 compensation point was associated with improved photosynthetic function.
This study investigated the effects of different shading levels on the growth, physiological characteristics of D. maackianum. Based on the results and discussions, several conclusions can be drawn.
Increased shading levels resulted in lower ambient temperatures and higher relative humidity, indicating that higher shading reduces direct sunlight exposure. This reduction in light leads to less temperature increase and decreased transpiration, thereby increasing relative humidity. All plants under full light conditions (control) perished due to excessive transpiration and water imbalance, which caused leaf desiccation.
In terms of morphological adaptations, the specific leaf area (LMA) was lowest in the 95% shading treatment, suggesting thinner and broader leaves. This is a common adaptation to low light conditions. However, there were no significant differences among treatment groups, suggesting a lack of significant morphological adaptation across different light levels.
Physiologically, chlorophyll content (a, b, and total) increased with higher shading levels, demonstrating typical shade-plant adaptations to maximize light absorption and photosynthetic efficiency under low light. The Fv/Fm and NPQ values showed no significant differences among treatment groups, indicating that the plants maintained photosystem II functionality and photoprotective mechanisms across varying light conditions.
The dark respiration rate (Rd) and light compensation point (LCP) decreased with increased shading, aligning with typical shade-tolerant plant responses. The 95% shading treatment exhibited the highest photosynthetic rates, light saturation points, and apparent quantum yields, indicating enhanced photosynthetic efficiency under low light conditions. Carboxylation efficiency showed no significant correlation with other traits, but CO2 compensation point and photorespiration rate had negative correlations with relative light intensity and positive correlations with chlorophyll content and maximum photosynthetic rate.
Analyzing the correlation between light intensity and physiological traits revealed negative correlations between relative light intensity and chlorophyll content, light compensation point, light saturation point, maximum photosynthetic rate, and apparent quantum yield. This suggests that D. maackianum optimizes photosynthesis under low light by increasing light absorption and utilization efficiency. Higher chlorophyll content is associated with increased photosynthetic activity, as evidenced by positive correlations with apparent quantum yield, maximum photosynthetic rate, CO2 compensation point, and photorespiration rate. D. maackianum exhibits significant physiological adaptations to optimize photosynthesis and growth under varying light conditions. These findings highlight the species’ ability to adjust its physiological processes and structural characteristics to cope with different shading environments, enhancing its survival and growth in shaded habitats.
