

서 론
우리나라의 기후는 온대성 기후대로 낙엽활엽수림은 전국적으로 분포하며, 상록활엽수림은 남해안 일대와 도서 지역에서 제한적으로 분포하고 있다(Lee et al., 2005). 난온대 상록활엽수림의 대표 식생은 붉가시나무, 참가시나무, 구실잣밤나무 및 후박나무로 보고된 바 있으며(Kim and Oh, 1997), 급속한 기온상승은 상록활엽수림에 분포와 생육에 직접적인 영향을 미친다(Yun et al., 2011). 기후변화 정부 간 협의체(IPCC: Intergovernmental Panel on Climate Change)의 제 6차 평가보고서에 따르면 2100년의 지구 평균기온은 1.5℃ 이상 상승으로 생물종 감소와 식생 변화에 따른 생태계의 위험성을 강조하고 있다(MaCarthy et al., 2001). 최근 특정 식물종의 생육특성을 파악하여 기후조건에 맞는 생육지의 잠재적 분포지역과 적정 생육지를 선정하여 기후변화에 대응할 수 있도록 도움을 준다(Horikawa et al., 2009; Tsuyama et al., 2008). 이러한 기술적 발전은 기후변화와 인위적인 훼손지 발생에 따른 단위면적 당 식물 공급 규모를 결정할 수 있다. 또한, 생태복원 계획을 수립할 때 참조생태계 관점에서 재배품종과 외래종보다는 훼손지 주변의 대표 식생을 파악하여 공급 체계를 강화할 수 있다.
현재 우리나라에서 조림과 훼손지역에 용기묘 공급은 약 20년 정도 진행되어 왔으며(Cho et al., 2012), 용기묘 대량생산을 위한 용기 용적률(Jin et al., 2015), 차광 여부 및 차광수준(Choi et al., 2012), 시비의 공급량(Choi et al., 2019; Ha et al., 2019; Kim et al., 2014) 등 객관적 자료가 뒷받침되고 있다. 또한, 시비는 비료와 토양의 특성을 정확히 분석해야 한다(Cho et al., 2012). 특히, 용기묘로 생산된 묘목이 훼손지에서 조림 이후 시비 처리가 되지 않기 때문에 주변 식생과의 초기 경쟁에서 우위를 점할 수 있도록 시비 효과에 대한 검증이 필요하다(Cho et al., 2012). 이를 위해서는 과량시비에 의한 묘목 및 토양의 피해가 없는 한도 안에서 수종별 적정 시비량 구명과 동시에 조림 묘목이 다른 하층식생보다 효율적으로 양분을 흡수할 수 있는 시비 기술이 구명되어야 한다.
본 연구에서는 양묘생산 과정에서 시비 처리를 실시하여 생육한 후박나무(Machilus thunbergii) 용기묘를 대상으로 간장, 근원경, 묘목품질지수, 광합성 반응 측정 등의 생장, 생리적 특성을 조사 및 분석하여 용기묘로 생산된 묘목의 효율적인 적정 시비량 및 시비 기술을 구명하고자 연구를 실시하였다. 또한 우수한 형질의 묘목 생산을 위한 대량 생산기술 개발 등의 필요한 정보를 얻고자 하였다.
재료 및 방법
공시수종 및 시비처리
실험에 사용된 재료는 전남 해남 일대에서 채취한 종자(북위 37° 45', 동경 127° 107')를 2021년 2월 해남농장에서 파종하여 생산된 후박나무 실생묘로서 한국농수산대학교 산림학과 온실에서 2022년 3월부터 9월까지 생육시키면서 실험을 진행하였다. 이때 임업시설양묘용 15구 플라스틱 용기(Shinill Science Inc., Gyeonggi, Korea; Table 1)에 피트모스, 펄라이트, 질석(1:1:1,v/v)의 상토를 채웠으며, 토양분석에 사용된 모든 방법은 농촌진흥청의 상토 표준분석법을 기준으로 실시하였다. 시비는 2022년 6월 1일부터 8월 31일까지 총 14주 동안, 주 2회씩 실시하였다. 수용성 비료인 MultiFeed 19(N P K=19 : 19 : 19. Haifa Chemicals, Israel)를 이용하여 규격별, 처리별 3 tray씩 12 tray에 총 360본의 용기묘에 20 L의 관수와 병행하여 500 ㎎/L, 1000 ㎎/L, 2000 ㎎/L 등의 4가지 시비처리를 하였다. 고온과 이상기온 현상에 의한 피해를 줄이기 위해 8∼9시 또는 17∼18시 사이에 관수와 시비를 진행하였다.
Table 1.
Chlorophyll and carotenoid contents of M. thunbergii saplings grown under different fertilization levels
| Treatment | Chl (㎎·g-1) |
Car (㎎·g-1) | Chl a/b | T Chl/Car | ||
| a | b | a + b | ||||
| Control | 3.38 nsz | 0.83 nsz | 4.21 nsz | 1.57 nsz | 4.61 nsz | 2.68 nsz |
| 500 ㎎/L | 2.27 | 0.50 | 2.78 | 1.06 | 4.64 | 2.63 |
| 1000 ㎎/L | 3.68 | 0.90 | 4.59 | 1.68 | 4.42 | 2.69 |
| 2000 ㎎/L | 3.69 | 0.74 | 4.44 | 1.69 | 5.22 | 2.57 |
생리특성조사
시비처리에 따른 광합성 반응은 생육상태가 건전하며 최상부에 위치한 잎을 대상으로 휴대용 광합성측정장치(Portable photosynthesis system, Li-6800, LI-COR Inc, USA)를 이용하여 측정하였다. 실험은 생육이 활발하게 이루어지는 6월 20일경에 6반복으로 진행하였다. 실험을 위해 휴대용 광합성 측정장치에 부착된 LED light source를 이용하여 챔버 내부 PPFD 1,200 μmol·m−2·s−1, CO2 농도 400 ± 2 ㎎/L, 광합성 측정기에 유입되는 공기의 유량 500 μmol·m−2·s−1, 온도 25 ± 2℃로 설정하여 챔버 내부로 안정적이게 공급하여 광포화 상태에서의 순광합성속도(net photosynthetic rate, A), 기공전도도(stomatal conductance, gs), 기공증산속도(stomatal transpiration rate, E)를 측정하였다. 또한, 순간증산효율(Instantaneous transpiration rate, ITE), 내재적 수분이용효율(Intrinsic water use efficiency, WUEi), 엽육세포간극내 CO2 농도 비율(intercellular CO2 concentration/ atmospheric CO2 concentration, Ci/Ca)을 산출하였다(Song et al., 2022).
2022년 9월 CO2 농도(0-1,400 μmol·m-2·s-1)에 의한 엽육세포내 CO2 반응 곡선(A-Ci)을 3반복으로 조사하여 최대카르복실화속도(maximum carboxylation rate, Vcmax) 및 최대전자전달속도(maximum electron transport rate, Jmax)를 산출하였다(Sharkey et al., 2007). 이때 측정 조건은 챔버로의 유입 공기유량을 600 μmol·s-1, 온도를 25±1 ℃로 유지하였다.
엽색소 함량 분석은 9월 초 최상단에 위치한 잎을 처리구당 5반복으로 채취하여 진행하였다. Hiscox와 Israelstarm (1978)의 방법에 따라 엽편 0.1 g을 10 mL의 DMSO (dimethyl sulfoxide) 용액이 들어있는 20 ㎖ 갈색 유리병에 넣어 60℃로 설정된 항온기에서 6시간 동안 추출하였다. 추출액은 UV-Visible spectrophotometer (UV/VIS Spectro-photometer, HP 8453, Hewlett-Packard, USA)를 이용하여 663 ㎚, 645 ㎚, 445 ㎚의 파장에서 흡광도를 측정하였다. 엽록소 a, b, 총 엽록소 함량 및 카로티노이드 함량을 산출하고, 엽록소 a/b 및 Total Chl/Car를 계산하였다(Arnon, 1949). 엽록소 형광 반응은 6월과 9월에 2회 측정하였으며, 엽록소 형광반응 측정기(Plant Efficiency Analyser, PEA, Hansatech Instrument Ltd., King’s Lynn, England)를 활용하였다. 형광반응 측정은 광합성 측정과 동일한 잎을 대상으로 leaf clip으로 광을 차단하여 측정 전 약 20분간 암 조건에 적응시킨 후 측정하였다. 측정은 3,500 μmol·m-2·s-1의 광량을 1초간 조사하였다. 따라서 50 μs (O단계), 300 μs (K단계), 2 ms (J단계), 30 ms (I단계), 500 ms (P단계)에서의 엽록소 형광밀도와 Fo/Fm, Fv/Fm, ABS/RC, DIo/RC, RC/CSo PIabs, SFIabs를 비교·분석하였다(Lee et al., 2021).
생장특성
시비처리에 따른 수고 및 근원경 생장을 조사하기 위하여 3월부터 8월까지 매월 1회, 처리구별 각 20 본씩 전자식 캘리퍼스와 스틸테이프를 이용하여 간장과 근원경을 반복 측정하였다. 특히, 불규칙한 근원의 모양에 따른 오차 값을 줄여주기 위해 가로와 세로 2 회 측정하여 평균값을 산출하였고, 양묘의 건전도를 판단할 수 있는 H/D [height (㎝)/root collar diameter(㎜)]을 산출하였다(Bayala et al., 2009). 또한 부위별 생장 차이를 알아보기 위해 광합성 실험 종료 후 10월 초에 각 처리구당 5개체씩 시료의 지상부(잎과 줄기)와 지하부(뿌리)로 구분하여 분석에 사용하였으며, 시료는 건조기(Dry-oven)에서 48시간 동안 80℃로 건조시킨 후, 측정 결과에 따라 T/R율(top/root ratio), 잎건중비(Leaf weight ratio; LWR), 줄기건중비(Shoot weight ratio; SWR), 뿌리건중비(Root weight ratio; RWR) 등 물질분배 비율을 산출하였다.
통계분석
실험 결과분석을 위한 통계는 SPSS Statistics program 19.0 (SPSS Inc., Chicago,. IL, USA)을 이용하였으며, 일원분산분석을 통해 처리구간에 차이를 5% 유의수준에서 검정하였고, 사후분석은 Tukey’s HSD로 진행하였다.
결과 및 고찰
광합성 특성
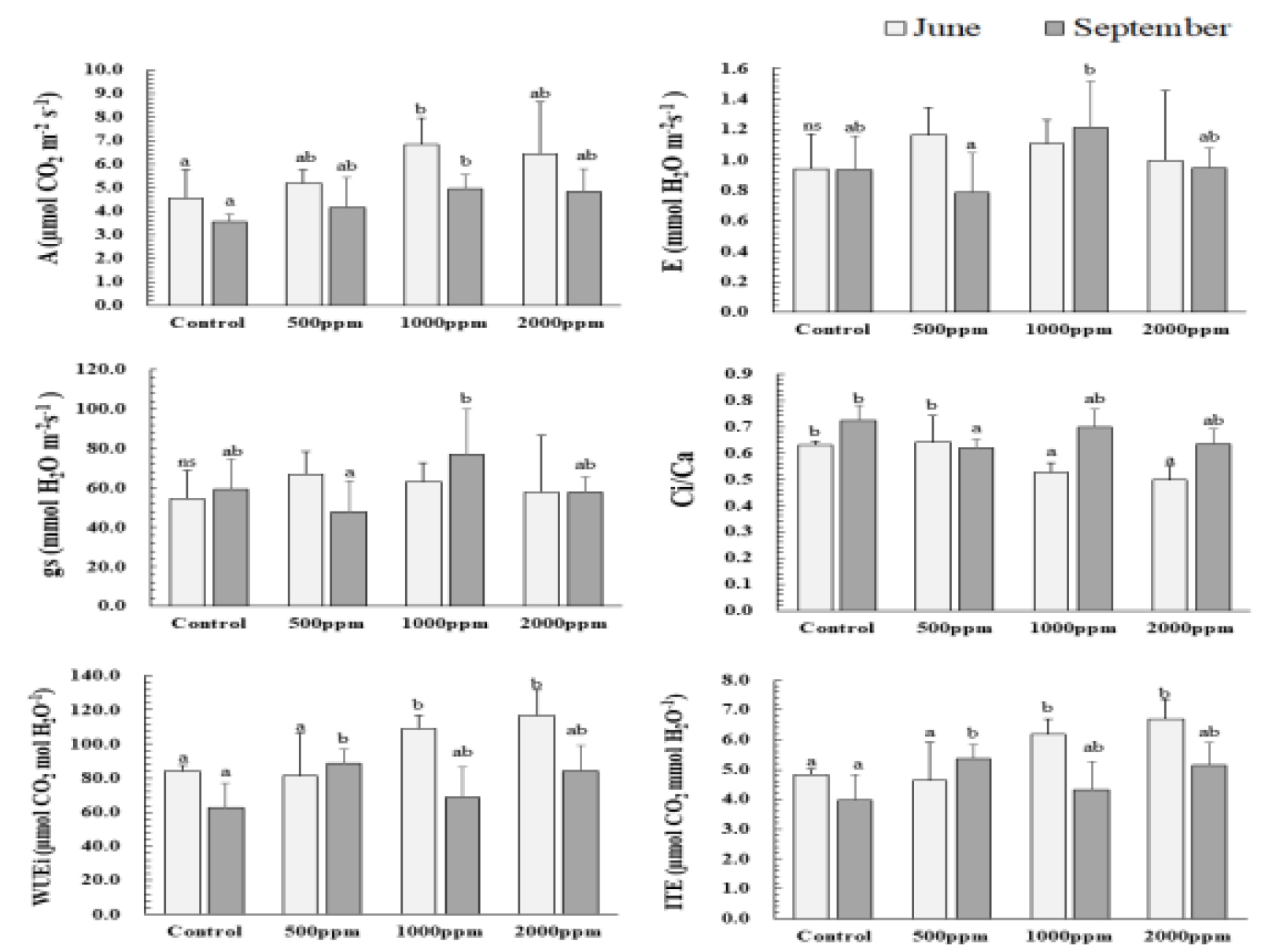
시비처리에 따른 순광합성속도(A)는 시비처리구(500, 1000, 2000 ㎎/L)에서 대조구와 비교하여 6월과 9월 각각 약 14-50%, 17-40% 높아 통계적인 유의성을 보였다(Fig. 1). 특히, 1000 ㎎/L은 6월에 6.9 μmol·CO2·m-2·s-1, 9월에 5.0 μmol·CO2·m-2·s-1로 가장 높은 A를 나타냈다. 기공증산속도(E)와 기공전도도(gs)는 6월 유의적인 결과가 나타나지 않았지만 9월, 1000 ㎎/L에서 가장 높은 경향을 보였다. 이는 다른 처리구들에 비해 상대적으로 잦은 기공개폐기작에 따른 순간증산효율(ITE)과 내재적 수분이용효율(WUEi)이 감소한 것을 볼 수 있었다. 이와 대조적으로 500 ㎎/L에서 gs와 E가 가장 낮아 모든 처리구들 중 가장 많은 기공을 닫고 있었으며, 수분이용효율 관련 지표들이 가장 높았던 것으로 나타났다.
대조구, 1000, 2000 ㎎/L에서의 높은 Ci/Ca는 엽육세포간극내 CO2 소비가 상대적으로 낮은 값을 보였지만 대조구와 2000 ㎎/L은 CO2의 소비가 원활하지 않았고 상대적으로 낮은 E, gs로 가스교환이 저조하였다. 따라서 A 역시 1000 ㎎/L에 비해 낮았던 것으로 여겨진다. 반면, 1000 ㎎/L은 높은 Ci/Ca를 유지하였지만, 높은 E, gs로 활발한 가스교환이 이루어지않아 생육이 왕성한 6월과 비교하면 낮은 값을 보였지만 9월에는 처리구들 중 가장 높은 A를 유지하였다. 한편, 500 ㎎/L은 대조구에 비해 A는 높게 유지하였지만, 유의적으로 가장 낮은 E, gs, Ci/Ca로 가스교환을 통한 CO2 흡수보다 이미 엽육세포간극내 축적되어 있는 CO2소비를 통해 수분손실을 최소화 하는 등의 효율적인 광합성 기작이 이루어진 것으로 판단된다.
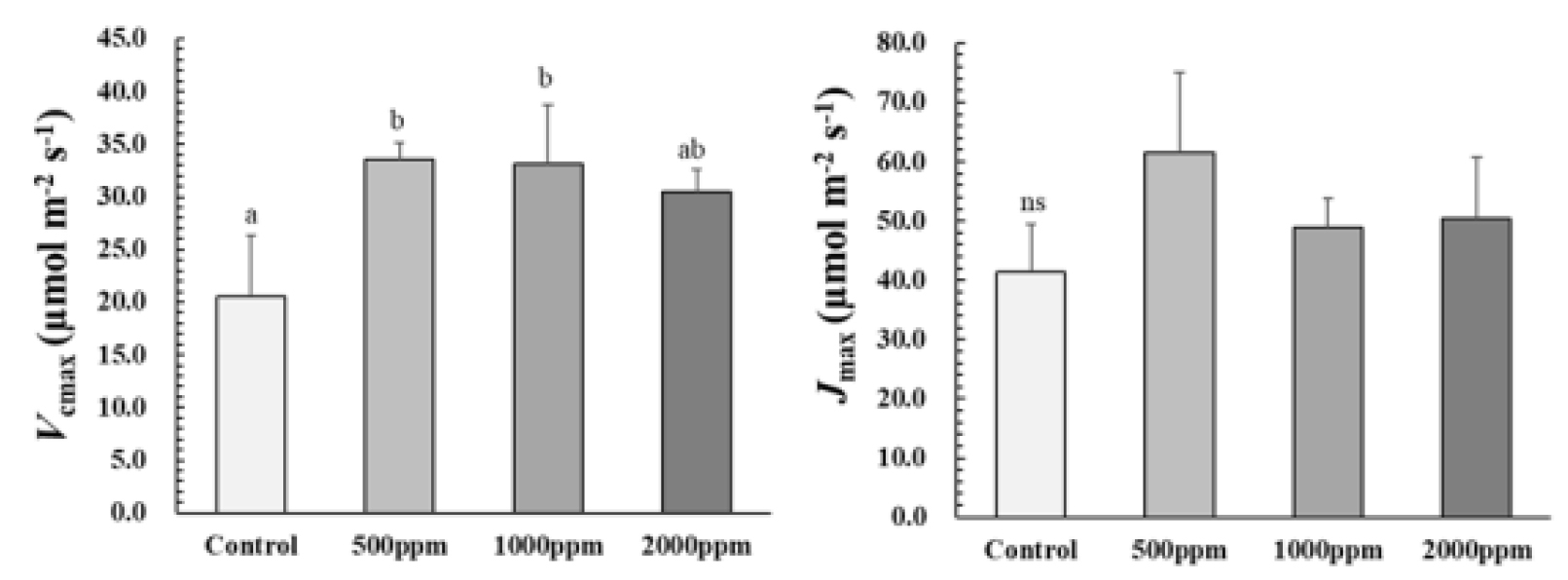
광합성 능력은 루비스코(ribulose-1,5-bisphosphate carboxylase/oxygenase; RuBisCO)의 활성에 따른 카르복실화율 및 리불로오스 이인산(ribulose bisphosphate; RuBP)의 재생속도간 균형에 의해 결정되며, RuBP의 재생속도는 전자전달 효율에 의해 제한된다(Ryu et al., 2014). A-Ci 곡선을 통해 산출한 후박나무의 최대 카르복실화 속도(maximum carboxylation rate; Vcmax)는 모든 시비처리구에서 대조구에 비해 약 1.5-1.6배 높았다. 특히, 2000 ㎎/L의 Vcmax는 30.5 μmol·m-2·s-1로 500, 1000 ㎎/L과 비교하여 약 8-10% 정도 낮은 것을 볼 수 있었다(Fig. 2). 이는 시비처리가 후박나무 용기묘의 광합성 기능 및 능력 향상에 도움을 주는 것을 알 수 있지만 2000 ㎎/L의 결과와 같이 과한 시비는 오히려 광합성 능력을 저하시킨 요인으로 나타났다. 최대 전자전달 속도(maximum electron transport ratel Jmax)는 시비처리구별 유의적인 차이가 나타나지 않았다(p > 0.05). 이는 시비처리가 광합성 능력을 증진시키는 데에는 양분을 광 수확 및 전자전달과 관련된 틸라코이드 단백질(Jmax와 연관)보다 RuBisCO(Vcmax와 연관)에 우선적으로 할당하여 나타난 결과로 여겨진다(Maier et al., 2008).
결과적으로 후박나무 용기묘는 대조구에 비해 시비처리구에서 순광합성속도(A), 순간증산효율(ITE), 내재적 수분이용효율(WUEi), 최대카르복실화속도(Vcmax) 등이 상대적으로 높았다. 500 ㎎/L은 가장 낮은 Ci/Ca와 양호한 A로 CO2 소비와 동시에 수분손실을 최소화 하여 효율적인 광합성 기작을 보였다. 또한, 1000 ㎎/L는 활발한 가스교환을 통해 가장 양호한 광합성 반응기작을 보였다. 상수리나무, 상동나무에서 역시 시비수준 1000 ㎎/L에서 가장 높은 A를 보였으며, A-Ci 곡선이 제시되었던 Eucalyptus pellita와 상동나무의 경우 1000 ㎎/L에서 높은 광합성능력이 유지된바 있다고 보고되었다(Cho et al., 2011; Eo et al., 2021; Kwon et al., 2009). 비교적 높은 시비농도를 가진 2000 ㎎/L에서 저조한 기공개폐기작으로 가스교환이 적었고 엽육세포간극내 CO2 소비 역시 원활하지 못해 전반적인 광합성 능력이 감소했던 것으로 판단된다. 이러한 경향은 E. pellita와 상동나무에서도 보였으며, 한편, Acacia mangium의 경우 2000 ㎎/L에서도 광합성 능력지표가 증가되었다고 보고된바 있어 수종별 적정 시비 수준의 중요성이 나타나는 결과였다(Cho et al., 2011; Eo et al., 2021). 따라서 광합성 반응기작 결과에 따른 후박나무 용기묘의 시비농도는 활발한 가스교환을 통해 비교적 높은 A, Vcmax 보였던 1000 ㎎/L과 기공개폐기작의 조절로 효율적인 방법으로 광합성 반응기작을 향상시켰던 500 ㎎/L이 적정한 것으로 나타났다.
엽색소 함량
시비처리에 따른 엽록소 및 카로티노이드 함량의 유의적인 차이는 없었지만(p > 0.05), 대체로 500 ㎎/L에서 다른 시비처리구들에 비해 다소 낮은 결과를 보였다. 이는 엽색소 함량의 감소가 수광 되는 빛 에너지과 명반응 감소로 이어졌다. 그러나 양분분배를 틸라코이드 단백질보다 RuBisCO에 우선적으로 할당하여 광합성 반응기작을 효율적으로 유지하였으며, 그 결과 500 ㎎/L은 비교적 높은 A와 Vcmax를 나타났다.
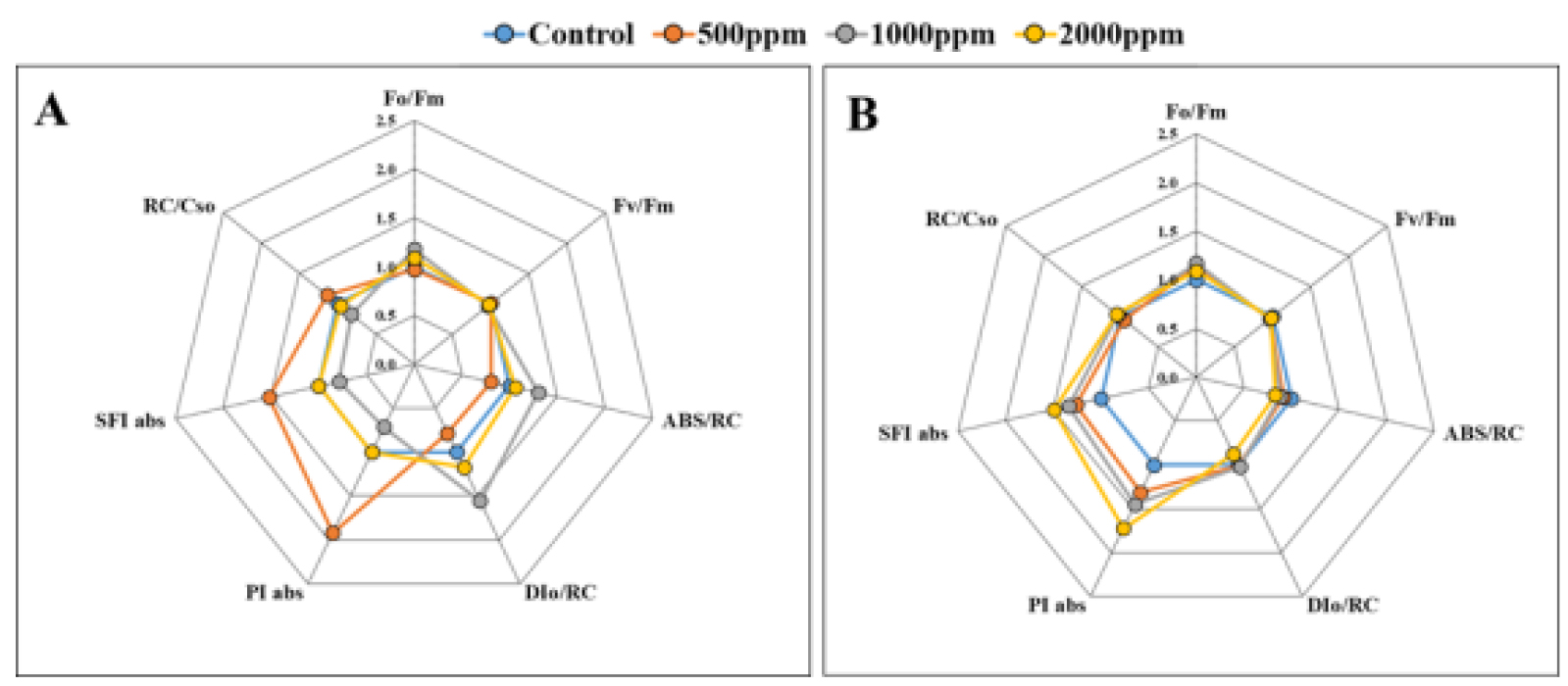
엽록소 a 형광반응 분석은 다양한 환경스트레스가 식물의 광합성 기구에 미치는 영향을 용이하고 정량적으로 제시할 수 있는 방법이다(Kalaji et al., 2017). 분석결과 6월과 9월 Fv/Fm, Fo/Fm, RC/CSo는 시비수준별 유의적인 차이를 보이지 않았다(p > 0.05 Fig. 3). 이러한 유의적 차이가 없는 결과는 상수리나무, 상동나무에서도 보고되었다(Kwon et al., 2009; Eo et al., 2021). 6월 500 ㎎/L이 대조구에 비해 PIabs와 SFIabs가 각각 약 1.9, 1.5배 증가하여 가장 양호하였다(Fig. 3A). 이는 500 ㎎/L이 엽색소 함량을 감소시켜 활력을 높였다고 여겨진다. 또한 1000, 2000 ㎎/L이 상대적으로 높은 농도의 시비조건으로 뿌리보다는 식물자체에서 받는 스트레스는 증가했음을 보여준다. 그러나 스트레스가 생리적 기능저하, 생장 감소로 이어지지 않는다는 점에서 효율적인 광합성 기구의 보호 기작이 이루어진 것으로 보인다. 특히, 1000, 2000 ㎎/L에서 반응중심 당 흡수된 빛에너지를 의미하는 ABS/RC와 열에 의해 소산되는 에너지를 나타내는 DIo/RC (Mathur et al., 2013)가 증가하는 것은 과잉여기에너지를 적극적으로 소산시키면서 전체적인 광합성 반응의 균형을 유도하고 있음을 알 수 있다. 9월에는 6월 1000, 2000 ㎎/L에서 증가되었던 ABS/RC 및 DIo/RC가 감소하여 대조구와 유사한 수준으로 보였다. 시비처리구에서 대조구에 비해 PIabs와 SFIabs가 약 1.3-1.7배 증가하여(Fig. 3B), 결과적으로 시비처리에 따라 광계 II에서 광계 I로 전달되는 에너지의 보존효율을 높이고, 광합성 기구의 활력 및 기능이 향상되는 반응을 보였다. 이에 따라 시비처리구에서 대조구에 비해 상대적으로 높은 순광합성속도(A) 및 최대카르복실화속도(Vcmax)를 보인 것으로 판단된다. 이러한 경향은 반상록관목 상동나무와 유사하였는데, PIabs가 시비처리구에서 대조구에 비해 약 3.1-3.4배 높았고, 이에 따라 시비처리구들에서 대조구에 비해 상대적으로 높은 A, Vcmax, Jmax를 유지한 것으로 보고된바 있다(Eo et al., 2021).
결과적으로 엽록소 및 카로티노이드 함량은 통계적인 유의성이 나타나지는 않았지만 500 ㎎/L의 엽색소 함량이 대조구를 비롯해 다른 시비처리구들에 비해 낮은 경향을 보였다. 이는 효율적인 광합성을 위한 대응기작으로 생각된다. 엽록소 a 형광반응의 경우 9월에 결과적으로 대조구에 비해 시비처리구들에서 에너지 보존효율이 약 1.3-1.7배 높아 광합성 기구의 활력 및 기능이 증진되었다고 볼 수 있다. 따라서 시비처리는 에너지 보존효율을 높이는 등 후박나무 용기묘의 엽록소 a 형광반응에 긍정적인 영향을 미쳤다고 판단된다.
생장량
근원경 생장은 500, 1000 ㎎/L이 유의하게 높았지만, 2000 ㎎/L은 대조구와 유사한 경향을 보여 상대적으로 낮은 근원경 생장을 보였다(Fig. 4). 간장 생장의 경우, 모든 시비처리구에서 대조구에 비해 약 3.0-3.5배 높았다. 따라서 전반적인 근원경 및 간장 생장은 1000, 500, 2000 ㎎/L, 그리고 대조구 순으로 나타났다. 상록활엽수인 굴거리나무, 가시나무, 붉가시나무, 종가시나무(Song et al., 2014; Kim et al., 2014)와 낙엽활엽수인 쉬나무(Choi et al., 2019)를 대상으로 한 시비 실험에서도 시비수준이 증가할수록 근원경과 간장 생장이 증가하여 후박나무와 유사한 결과를 보인 것으로 나타났다. 한편, Imo and Timmer (1999)는 수종별 양분 요구도가 상이하여 오히려 과시비로 인한 피해가 발생될 수 있다고 보고하였는데, Mutifeed 19를 2000 ㎎/L 시비한 후박나무의 경우 근원경 생장이 500, 1000 ㎎/L에 비해 낮았고, 통계적인 차이는 없었지만 간장 역시 상대적으로 다른 시비처리구들에 비해 낮은 결과를 보여 과시비로 인한 생육저하로 판단된다.
H/D율은 근원경 및 간장과 유사한 경향을 보여 대조구에서 3.4 ㎝·㎜-1로 가장 낮았으며, 시비수준이 증가할수록 높아지는 경향을 보였고(Fig. 4), 이러한 경향은 가시나무(Sung et al., 2020), 쉬나무(Choi et al., 2019), 굴거리나무(Song et al., 2014)에서 역시 유사한 경향을 보였다. 한편, 활엽수 노지묘의 바람직한 H/D율은 7.0 이하로 보고된 바 있는데(Johnson, 1996), 500-2000 ㎎/L의 농도로 시비처리 된 후박나무 용기묘의 경우 H/D율이 5.5-6.3의 범위로 나타난 것으로 미루어 보아 건전한 묘목인 것으로 판단된다.
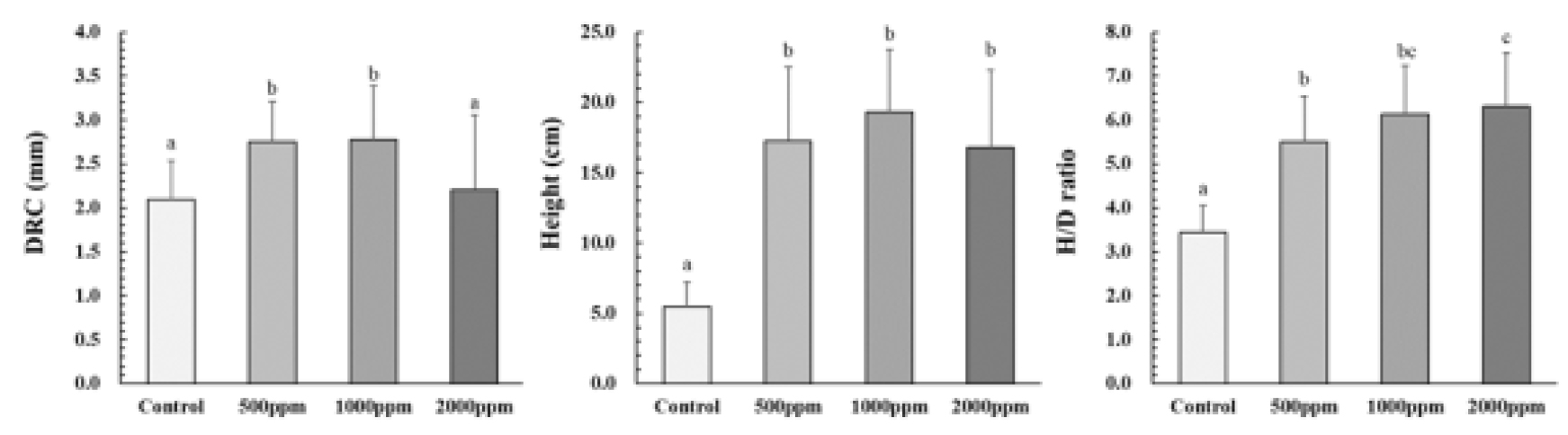
Fig. 4.
Effects on the diameter at root collar (DRC), Height, and H/D ratio of M. thunbergii under different fertilization levels. The data indicates means ± SD (n = 30). Different letters indicate a significant difference at p < 0.05 and ns meaning non-significance among different level of fertilization within the same season.
묘목의 건물생산량은 조림지 식재 후 활착률에 영향을 미치는 요소이며(Ritchie, 1984), 시비수준에 따른 후박나무의 전반적인 물질생산량은 시비처리구에서 대조구에 비해 유의적으로 높은 것을 볼 수 있었다(Table 2). 전체 물질생산량은 500-2000 ㎎/L에서 대조구에 비해 약 2.3-3.1배 높았으며, 유의성이 검정되지 않은 지하부(p > 0.05)보다 지상부 생장(잎, 줄기)의 차이로 인한 결과로 판단된다.
T/R율은 대조구에서 가장 낮은 1.17 g·g-1로 시비처리구에 비해 약 2.4-3.6배 낮은 수준이었고, 이는 충분한 양분 조건의 시비처리구에서 활발한 광합성 활동을 통해 전반적인 생장이 증가하여 나타난 결과로 판단된다(Timmer and Armstrong, 1987). 굴거리나무(Song et al., 2014)와 가시나무(Sung et al., 2020) 역시 시비수준이 증가함에 따라 T/R율이 증가하여 후박나무와 유사한 경향을 보였다. 특히, 1000 ㎎/L의 T/R율은 유의적인 차이는 아니지만 500, 2000 ㎎/L에 비해 다소 낮아, 지하부의 생장이 두 처리구에 비해 상대적으로 양호했던 것으로 여겨진다. Oh (1982)는 우리나라 묘목의 T/R율의 범위가 2.5-3.0 g·g-1 정도면 건전한 묘목이라고 보고하였는데, 후박나무 1000 ㎎/L의 T/R율이 2.81 g·g-1로 나타나 건전한 묘목의 형태를 보였다고 판단된다. 광합성 산물의 물질분배율을 보여주는 잎건중비(LWR), 줄기건중비(SWR), 뿌리건중비(RWR) 결과는 Table 2와 같이 나타났다. 시비처리구 지상부의 물질분배율 LWR과 SWR은 대조구에 비해 각각 약 46-60%, 22-33% 높았지만 RWR은 이와 상반되는 결과를 보여 대조구에서 0.48 g·g-1로 시비처리구에 비해 약 1.4-2.4배 높아 가장 높은 RWR을 보였다. 선행연구에서 보고된 바와 같이 시비수준이 증가할수록 RWR이 감소하는 결과를 얻었으며(Choi et al., 2019; Reynolds and Antonio, 1996; Schlichting, 1986; Sung et al., 2020), 이는 시비수준에 따라 생육환경 조건이 달라져 식물의 생장 반응이 조절되어 RWR이 감소한 것으로 사료되며(Gleeson, 1993; Hilbert, 1990), 시비처리가 지하부보다 상대적으로 지상부에 더 많은 영향을 미치는 것으로 생각된다(Reynolds and Antonio, 1996).
Table 2.
Effects of fertilization levels on the dry mass, T/R ratio, LWR, SWR, and RWR with M. thunbergii saplings.
| Treatment | Dry mass production(g) |
T/R ratio (g·g-1) |
LWR (g·g-1) |
SWR (g·g-1) |
RWR (g·g-1) | |||
| Leaves | Shoot | Root | Total | |||||
| Control | 1.31 az | 0.69 a | 1.89 ns | 3.89 a | 1.17 a | 0.35 a | 0.18 a | 0.48 b |
| 500 ㎎/L | 4.74 b | 2.04 b | 2.13 | 8.91 b | 3.39 b | 0.54 b | 0.23 ab | 0.24 a |
| 1000 ㎎/L | 5.70 ab | 2.41 ab | 3.01 | 11.12 b | 2.81 ab | 0.51 b | 0.22 ab | 0.27 a |
| 2000 ㎎/L | 6.69 b | 2.78 b | 2.54 | 12.01 b | 4.23 b | 0.56 b | 0.24 b | 0.20 a |
적 요
후박나무 묘목의 효율적인 생산을 위해 시비농도에 따른 간장, 근원경, 묘목품질지수, 광합성 반응 측정으로 생장과 생리적 특성을 조사하고 적정 용기묘 생산에 적합한 시비량을 확인하였다. 광합성 특성은 대조구에 비해 시비처리구에서 순광합성속도(A), 순간증산효율(ITE), 내재적 수분이용효율(WUEi), 최대카르복실화속도(Vcmax) 등 높았다. 특히, 시비 농도는 활발한 가스교환을 통해 비교적 높은 A, Vcmax 보였던 1000 ㎎/L과 기공개폐 기작의 조절로 광합성 반응기작을 향상시킨 500 ㎎/L가 적정 수준의 시비로 실험 결과를 보였다.
생장량 또한 대조구보다 시비처리구에서 묘목의 품질지수 등 통계적으로 높은 것으로 나타났다. 특히, 1000 ㎎/L은 근원경, 간장이 처리구 중 가장 큰 특징을 보였으며, 잎, 줄기, 전체 건중량, 역시 다른 처리구들에 비해 통계적으로 높았고, 묘목의 품질을 나타내는 H/D율 및 T/R율도 건전한 수준인 것을 볼 수 있었다. 500 ㎎/L 역시 양호한 생육 특성을 보여 경제성을 고려한다면 한 가지 선택지가 될 수 있다고 여겨진다. 그러나 2000 ㎎/L의 경우, 근원경의 감소로 H/D율은 높아졌고, 지상부에 비해 지하부로의 물질분배가 저조하여 T/R율 역시 증가하는 형태적 특성과 비용 저감을 위한 경제성을 고려하였을 때 2000 ㎎/L의 시비농도는 과하다고 판단된다. 따라서 후박나무 용기묘의 시비는 500 ㎎/L 혹은 1000 ㎎/L이 경제적으로나 식물의 형태적으로 가장 이상적인 시비량이라고 판단된다.
