

서 언
다양한 유전자원의 확보와 평가는 다수확 및 고품질과 같은 우수한 형질을 발굴하고 새로운 교배 조합을 설계하는 가장 핵심적인 기반이 된다. 이를 체계적으로 평가하고 적극적으로 활용하는 것은 변화하는 재배 환경에 적응하면서도 육종 목적에 부합하는 최적의 신품종을 성공적으로 개발하기 위한 필수 조건이다. 최근 유전자원의 효율적 활용을 위해 국내 종자은행 보존 자원의 다양성을 분석한 연구가 보고되었고(Yu et al., 2025), 유채 또는 메밀을 대상으로 농업적 특성과 유붕 성분 조성 간의 상호관계 및 군집분석을 진행하여 유용한 육종 소재 정보를 제공한 사례도 있다(Cho et al., 2025, Kim et al., 2020).
트리티케일(x Triticosecale Wittmack)은 밀의 우수한 품질과 호밀의 환경적응성(한냉해, 가뭄, 염해, 병충해 저항성 등)을 결합하여 육성된 작물이다. 1875년 Stephen Wilson과 1891년 Rimpau에 의해 트리티케일 개발에 성공한 이후, 미숙배 배양과 콜히친 처리에 의한 염색체 배가 기술 등을 통해 안정적인 1차 트리티케일 개발(속간 교잡) 기술이 확립되어 오다가 1960년대부터는 불임 혹은 임성이 떨어지는 트리티케일을 밀과 교배하거나(Sisodia and McGinnis, 1970a) 기존 트리티케일과 교배하여(Sisodia and McGinnis, 1970b) 새로운 유전적 변이를 도입함으로써 형질이 안정적으로 유전되는 2차 트리티케일(종내 계통 교배) 계통을 육성하는 등 본격적인 실용 식물육종의 대상이 되었다(Briggle, 1969; Gupta and Priyadarshan, 1982; Mergoum et al., 2019). 1964년 국제옥수수밀연구소(CIMMYT)에서 트리티케일 육종을 시작하였고(Zillinsky, 1973), 1967년 CIMMYT에서는 곡립이 잘 맺히는 ‘고임성’, ‘곡립위축현상’의 개선, 낮의 길이 변화에 크게 영향을 받지 않아 재배지역의 폭을 넓힐 수 있는 ‘일장둔감형’, 그리고 키를 작게 하여 잘 쓰러지지 않게 하는 ‘반왜성’ 유전자가 도입된 아르마딜로(Armadillo) 계통을 개발하였다(Mergoum et al., 2019; Zillinsky, 1973). 1968년 헝가리에서 No. 57과 No. 64 품종, 1969년 캐나다에서 Rosner 품종이 개발되는 등 1970년대 이후 상업적 재배가 시작되었고(Gupta and Priyadarshan, 1982), 최근에는 폴란드, 벨라루스, 독일 등 유럽을 중심으로 재배되며, 2024년 기준으로 전 세계 44개국에서 347만 ha 규모로 재배되어 연간 1,277만톤이 생산되고 있다(FAO, 2024).
트리티케일은 사료, 식용 및 산업용 등 다양한 용도로 활용되고 있으며(Müntzing, 1979; NRC, 1989), 주로 조사료 및 알곡사료 형태로 축산업에 이용된다(Mergoum et al., 2019; Royo et al., 1993). 조사료용으로는 수분을 60 ~ 70%로 함유한 상태로 발효시켜 저장하는 ‘사일리지’, 수분을 말린 ‘건초’, 푸를 때 베어서 바로 먹이는 ‘청예(풋베기)’, 그리고 가축을 풀어 직접 뜯어 먹게 하는 ‘방목’ 등으로 다양하게 활용되고 있고(Derycke et al., 2018), 국내에서는 주로 사일리지 형태로 이용되고 있다(Kim et al., 2022; Lee and Choi, 2017). 지속농업을 위한 경지관리를 위해 트리티케일을 피복작물로 이용 후 수확하여 사일리지를 제조하여 젖소 사양에서 옥수수 사일리지 일부를 대체한 사례도 보고되었고(Harper et al., 2017), 트리티케일을 활용한 이모작이 작물 병해를 경제적으로 관리 가능한 수준으로 유지하는 데 효과적임을 보고하였다(Sumner et al., 1995). 곡실사료 용도로는 트리티케일 곡실은 전분과 건물 함량이 높아 소화율이 우수하며, 필수 아미노산 함량이 높아 돼지, 가금류, 반추동물(소, 양) 배합사료로 적합하다고 하였다(Mergoum et al., 2019). 비육우 사양에서 트리티케일 증류건조박은 일부 조사료 대체가 가능하며(Wierenga et al., 2010), 육계사료에서 밀, 옥수수 대체원으로 활용한 연구도 보고되었다(Osek et al., 2010). 식용 용도로는 제빵용 트리티케일 육종(Ghendov-Mosanu et al., 2024; Lukaszeski et al., 2006) 뿐만 아니라 제빵에 트리티케일 가루를 밀가루와 최대 50%까지 혼합이 가능하였다(Naeem et al., 2002). 비스킷 제조시에도 밀가루를 50% 대체하고(Yaseen et al., 2007), 파스타 건면 제조용으로도 활용 가능하며(Cervini et al., 2024), 항산화 성분이 풍부한 기능성 식품 원료로서 큰 잠재력이 있음이 보고되었다(Kaszuba et al., 2021). 재생에너지 생산이라는 측면에서는 트리티케일이 바이오 에탄올 및 가스 생산을 위한 에너지 작물로도 주목받고 있다(Cantale et al., 2016). 캐나다 대평원에서 곡물과 바이오매스를 밀보다 안정적으로 높은 수량을 내는 트리티케일 유전자원이 선발되었고(Goyal et al., 2011), 트리티케일 곡물을 이용한 에탄올 생산과 수율이 보고되었으며(Klikocka et al., 2019; Wang et al., 1997), 트리티케일 짚을 이용한 에탄올과 바이오가스 생산 수율이 보고되었다(Łukajtis et al., 2018; Vasiljevic et al., 2021). 섬유 자원으로서 트리티케일은 짚 수량이 높고, 짚의 보수력이 커 가축 깔짚 및 바이오소재 잠재력이 크다고 제시된 바 있다(Derycke et al., 2018).
우리나라에서 트리티케일 연구는 1970년대 농촌진흥청 국립식량과학원과 멕시코에 있는 국제옥수수밀연구소(CIMMYT)와의 공동연구를 추진함으로써 시작되었다(Kim et al., 2020). 현재까지 국립식량과학원에서 ‘조성’, ‘신영’을 비롯한 12개 품종을 육성하여 보급하고 있으며(NICS, 2025), 벼를 수확한 후 이어서 겨울철에 트리티케일을 재배하는 논 이모작 재배에 적합한 숙기가 빠르고 조사료 생산성이 우수한 특성을 가진 품종을 지속적으로 개발해 오고 있다. 그러나 조사료용 품종개발에 있어 안전재배지역을 중북부로 확대하기 위해 내한성을 강화시킨 조생 품종개발이 요구됨에도 불구하고(Kim et al., 2020), 육종의 기본 바탕이 되는 유전자원의 내한성과 농업 형질에 대한 평가와 형질간 유연관계 분석이 미흡한 실정이다. 본 연구에서는 트리티케일 유전자원 형질의 다양성을 조사하고, 각 형질 간 상호 관계를 분석하여 유사한 자원 간 군집화(clustering)를 통해 집단을 대표하는 특이적 형질을 도출하여 조사료용 내한성 트리티케일 품종개발을 위한 교배집단 선발의 기초자료로 활용하고자 하였다.
재료 및 방법
실험재료
본 연구에는 국립식량과학원이 보유한 253점의 트리티케일 유전자원을 이용하였다. 총 17개국에서 수집되었으며, 주요 수집국으로는 미국 96 자원(37.9%), 한국 55 자원(21.7%), 러시아 37 자원(14.6%)이었고, 멕시코에 소재한 국제옥수수밀연구소(CIMMYT)로부터 도입된 27 자원(10.7%)도 포함되었다. 이 외 캐나다, 프랑스, 스페인 등 13개 국가에서 수집된 자원들이 이용되었다(Supplementary Table S1). 유전자원의 원산지 지역별 그룹화(origin regional group)는 각 국가의 지정학적 위치와 농업기후적 유사성을 고려하여 동아시아(한국, 중국, 일본), 북아메리카(미국, 캐나다), 라틴아메리카(멕시코),북·동유럽(러시아, 스웨덴, 핀란드, 폴란드, 우크라이나, 불가리아, 헝가리), 서유럽(프랑스, 스페인), 그리고 오세아니아·아프리카(호주, 남아공) 등 총 6개 지역으로 분류하였다(Table 1). 유전자원 중 TG125는 월동 중에 전 개체가 고사하여 월동 후 형질 조사에서는 제외되었다.
Table 1.
Origin grouping of the 253 triticale genetic resources.
재배방법 및 농업 형질 평가
본 연구의 유전자원 재배 및 형질 평가는 2024년 10월부터 2025년 7월까지 국립식량과학원 중북부작물연구센터(수원, 37°16′ N 126°59′ E, 해발 31 m) 시험포장에서 수행되었다. 파종은 2024년 10월 17일에 실시하였으며, 시험구 배치는 자원당 1열씩 0.4 m × 2.0 m (휴간 × 휴장) 간격으로 조성하였다. 자원 간 비교를 위한 대비품종으로는 ‘신영’과 ‘조성’을 선정하여 매 20자원마다 배치하였다. 시비량은 10a당 N-P2O5-K2O를 12-9-7 ㎏으로 시용하였으며, 질소질 비료는 기비(2024년 10월)와 월동 후 생육재생기(2025년 3월 상순)에 각각 50%씩 분시하였다. 농업 형질 특성 조사는 농업과학기술 연구조사분석기준(RDA, 2012)과 국립종자원 트리티케일 특성조사기준(KSVS, 2024)에 준하여 수행하였다. 내한성 평가는 맥류 연구조사분석기준에 따라 월동 후 생육재생기인 3월 상순에 잎마름 비율이 20% 이하는 매우 강(1), 21∼40%는 강(3), 41 ∼ 60%는 중간(5), 61 ∼ 80%는 약(7), 80% 이상은 매우 약(9)으로 평가하였다. 양적형질은 자원당 10개체를 임의로 선택하여 내한성(WH, Winter Hardiness), 개화소요일수(DH, Days to Heading time), 초장(PH, Plant Height), 간장(CL, Culm Length), 수장(SL, Spike Length), 까락길이(AL, Awn Length), 지엽길이(FL, Flag leaf Length), 지엽너비(FW, Flag leaf Width), 천립중(TW, Thousand-grain Weight) 등을 조사하였다. 질적형질은 엽색 농담(Leaf color Intensity), 월동직전 생장정도(Degree of growth before winter), 월동직후 초형(Post-winter growth habit), 지엽 직립빈도(Frequency of recurved flag leaves), 엽이 안토시아닌 착색정도(Auricle anthocyanin coloration), 까락 안토시아닌 착색정도(Awn anthocyanin coloration), 꽃밥 안토시아닌 착색정도(Anther anthocyanin coloration), 지엽엽초 백분체농담(Glaucosity of Flag leaf Sheath), 지엽엽신 뒷면 백분체농담(Glaucosity of flag leaf lower side), 이삭 백분체농담(Spike glaucosity) 줄기 이삭목 털밀도(Density of hairs on neck), 이삭 까락 분포(Awns distribution of spike), 그리고 이삭 자세(Spike attitude) 등을 조사하였다.
통계분석
본 연구의 통계분석은 통계프로그램 R 소프트웨어(www.R-project.org, v.4.4.1., Vienna, Austria)를 사용하였다. 상관관계 분석은 피어슨 상관계수(Pearson Correlation Coefficient, PCC)로 유의성을 확인하였고, 유전자원의 형질 요인 간 주성분분석(Principal Component Analysis, PCA)을 수행하여 형질별 분포와 주성분의 기여도를 분석하였다. 유전자원 간의 유연관계 분석을 위해 형질의 표준화된 값을 이용하였으며, Euclidean 거리 척도로 Ward’s method를 사용하여 계층적 군집분석(Hierarchical Clustering Analysis)을 수행하였다.
결 과
트리티케일 유전자원의 형질 변이
유전자원의 원산지 그룹별 질적인 형질들의 분포 비율은 Table 2와 같다. 내한성 특성에서 전체 유전자원에서 ‘강함’ 이상의 비율이 51.8%로 나타나 내한성 트리티케일 개발에 활용될 수 있는 자원이 많이 있는 것으로 나타났다. 원산지 지역별로 보면 동아시아, 북아메리카, 서유럽 원산 유전자원에서 내한성 ‘강함’ 이상의 비율이 54% 이상으로 과반 이상의 비율을 차지했다. 서유럽 원산 자원의 87.5%는 잎 색이 농녹색인 반면, 북미 자원의 72.0%는 일반적인 녹색이었다. 지엽 만곡 빈도는 전체 유전자원의 절반 이상인 54.8%가 ‘매우 낮음’으로 나타나 지엽이 직립하는 특성이 우세하였다. 엽이의 안토시아닌 착색 정도는 전체 자원의 48.4%에서 ‘진함’을 보였는데 CIMMYT, 서유럽, 오세아니아·아프리카 원산 자원에서는 ‘진함’이 63 ~ 75%로 더 많이 관찰되었다. 수술 꽃밥의 안토시아닌 착색 정도는 전체 자원의 85.7%에서 안토시아닌 착색이 ‘없거나 약함’으로 나타났다. 이삭 백분체 농담은 ‘중간’ 정도의 백분체를 가진 자원이 51.2%로 가장 흔했으며, 동아시아 원산 자원은 이삭 백분체가 ‘많음’ 비율이 47.4%, ‘중간’ 비율이 43.9%로 비슷하게 분포했다. 엽초 백분체 농담에서는 전체 자원의 69.4%가 ‘매우 진함’을 보여 전반적으로 트리티케일 엽초 부위에 백분체가 많음을 알 수 있었고, 잎 뒷면 백분체 정도는 엽초에 비해 상대적으로 엷어 ‘중간’ 33.7%, ‘진함’ 35.7% 수준에 주로 분포하였다. 이삭 까락 분포는 ‘전체 분포’ 비율이 동아시아 원산 자원에서는 96.5%, CIMMYT 원산 자원에서는 96.3%로 대부분을 차지하였고, 북·동유럽 원산 자원은 ‘이삭 상부에만’ 까락 분포 비율이 28.9%로 타지역 원산지보다 상대적으로 높았다. 이삭 자세는 동아시아 자원의 79.0%가 ‘숙임형’이고, 1.8%가 ‘직립형’인 반면, 북·동유럽 자원은 6.7%가 ‘숙임형’, 55.6%가 ‘직립형’으로 뚜렷한 차이를 보였다. 월동 전 생장정도는 전체 자원 중 ‘약간 길음’ 29.3%, ‘길음’ 51.0%를 차지하여 트리티케일의 초기생장이 전반적으로 빠른 편임을 알 수 있었고, 이 중 동아시아, 북아메리카, 그리고 오세아니아·아프리카 원산지 자원들이 다른 지역 원산지 자원들 보다 초기생장이 빠른 자원 비율이 높았다. 월동 후 초형은 동아시아 원산지 자원의 75.4%가 ‘반직립’ 초형을 나타냈고, CIMMYT 원산지 자원은 77.8%가 ‘중간’ 초형을 보였다.
Table 2.
Percentage of each class for qualitatively-scored characters across origin groups and the entire triticale collection.
| Characters | Class | East Asia |
North America |
Latin America (CIMMYT) |
Northern & Eastern Europe |
Western Europe |
Oceania & Africa | Total |
| (n=57) | (n=108) | (n=27) | (n=45) | (n=8) | (n=8) | (n=253) | ||
|
Winter hardinessz | Very strong | 19.3 | 19.4 | 7.4 | 20.0 | 37.5 | 0 | 18.2 |
| Strong | 35.1 | 35.2 | 40.7 | 26.7 | 50.0 | 0 | 33.6 | |
| Medium | 24.6 | 32.4 | 29.6 | 28.9 | 12.5 | 75.0 | 30.4 | |
| Weak | 17.5 | 10.2 | 18.5 | 13.3 | 0 | 0 | 12.7 | |
| Very weak | 3.5 | 2.8 | 3.7 | 11.1 | 0 | 25.0 | 5.1 | |
| Leaf color | Dark green | 31.6 | 15.9 | 22.2 | 20.0 | 87.5 | 12.5 | 20.2 |
| Green | 50.9 | 72.0 | 56.6 | 51.1 | 12.5 | 50.0 | 61.5 | |
| Light green | 17.5 | 12.2 | 22.2 | 28.9 | 0 | 37.5 | 18.3 | |
|
Frequency of recurved flag leaves | Very low | 43.9 | 68.2 | 37.0 | 46.7 | 37.5 | 75.0 | 54.8 |
| Low | 36.8 | 21.5 | 22.2 | 35.6 | 25.0 | 12.5 | 27.4 | |
| Medium | 12.3 | 3.7 | 7.4 | 4.4 | 25.0 | 0 | 6.8 | |
| High | 3.5 | 6.5 | 14.8 | 6.7 | 12.5 | 12.5 | 7.0 | |
| Very high | 3.5 | 0 | 18.5 | 6.7 | 0 | 0 | 4.0 | |
|
Auricle anthocyanin coloration | Absent | 49.1 | 27.1 | 25.9 | 20.0 | 12.5 | 12.5 | 29.8 |
| Medium | 10.5 | 26.2 | 11.1 | 33.3 | 25.0 | 12.5 | 21.8 | |
| Strong | 40.4 | 46.7 | 63.0 | 46.7 | 62.5 | 75.0 | 48.4 | |
|
Awn anthocyanin coloration | Very weak | 19.3 | 67.3 | 51.9 | 82.2 | 50.0 | 50.0 | 56.4 |
| Weak | 56.1 | 28.0 | 40.7 | 11.1 | 37.5 | 50.0 | 33.7 | |
| Medium | 22.8 | 3.7 | 7.4 | 4.4 | 12.5 | 0 | 9.1 | |
| Strong | 1.8 | 0.9 | 0 | 0 | 0 | 0 | 0.8 | |
|
Anther anthocyanin coloration |
Absent or weak | 86.0 | 84.1 | 100 | 80.0 | 87.5 | 87.5 | 85.7 |
| Medium | 1.8 | 10.3 | 0 | 15.6 | 12.5 | 12.5 | 7.9 | |
| Strong | 12.3 | 5.6 | 0 | 4.4 | 0 | 0 | 6.4 | |
|
Spike glaucosity | Very strong | 5.3 | 0 | 0 | 0 | 0 | 0 | 1.2 |
| Strong | 47.4 | 12.2 | 11.1 | 22.2 | 25.0 | 25.0 | 22.6 | |
| Medium | 43.9 | 56.1 | 74.1 | 35.6 | 50.0 | 50.0 | 51.2 | |
| Weak | 1.8 | 24.3 | 14.8 | 28.9 | 25.0 | 12.5 | 18.7 | |
| Very weak | 1.8 | 7.5 | 0 | 13.3 | 0 | 12.5 | 6.4 | |
|
Glaucosity of sheath | Very strong | 80.7 | 64.5 | 77.8 | 62.2 | 75.0 | 62.5 | 69.4 |
| Strong | 15.8 | 22.4 | 14.8 | 33.3 | 25.0 | 25.0 | 22.2 | |
| Medium | 1.8 | 10.3 | 7.4 | 4.4 | 0 | 12.5 | 6.8 | |
| Weak | 1.8 | 1.9 | 0 | 0 | 0 | 0 | 1.2 | |
| Very weak | 0 | 0.9 | 0 | 0 | 0 | 0 | 0.4 | |
|
Glaucosity of lower side of blade | Very strong | 0 | 8.4 | 7.4 | 20.0 | 12.5 | 12.5 | 8.7 |
| Strong | 35.1 | 36.5 | 25.9 | 40.0 | 37.5 | 37.5 | 35.7 | |
| Medium | 52.6 | 26.2 | 48.2 | 20.0 | 50.0 | 12.5 | 33.7 | |
| Weak | 10.5 | 21.5 | 18.5 | 17.8 | 0 | 25.0 | 17.5 | |
| Very weak | 1.8 | 7.5 | 0 | 2.2 | 0 | 12.5 | 4.4 | |
|
Density of hairs on neck |
Absent or very sparse | 7.0 | 23.4 | 7.4 | 37.8 | 37.5 | 50.0 | 21.8 |
| Sparse | 15.8 | 10.3 | 14.8 | 17.8 | 12.5 | 0.0 | 13.1 | |
| Medium | 12.3 | 11.2 | 11.1 | 17.8 | 12.5 | 37.5 | 13.5 | |
| Dense | 31.6 | 35.5 | 51.9 | 22.2 | 25.0 | 0.0 | 32.5 | |
| Very dense | 33.3 | 19.6 | 14.8 | 4.4 | 12.5 | 12.5 | 19.1 | |
|
Awn distribution on spike | Tip awned | 0 | 3.7 | 3.7 | 28.9 | 0 | 0 | 7.1 |
| Half awned | 3.5 | 5.6 | 0.0 | 20.0 | 25.0 | 100 | 7.5 | |
| Fully awned | 96.5 | 87.9 | 96.3 | 51.1 | 75.0 | 0 | 84.1 | |
| Awns absent | 0 | 2.8 | 0 | 0 | 0 | 0 | 1.2 | |
| Spike attitude | Erect | 1.8 | 26.2 | 7.4 | 55.6 | 0 | 25.0 | 23.0 |
| Horizontal | 19.3 | 36.5 | 25.9 | 37.8 | 50.0 | 50.0 | 32.5 | |
| Drooping | 79.0 | 37.4 | 66.7 | 6.7 | 50.0 | 25.0 | 44.4 | |
|
Degree of growth before winter | Long | 0 | 20.4 | 0 | 0 | 0 | 12.5 | 29.3 |
|
Moderately long | 57.9 | 51.9 | 29.6 | 15.6 | 37.5 | 50.0 | 51.0 | |
| Medium | 42.1 | 25.9 | 59.3 | 55.6 | 50.0 | 25.0 | 17.4 | |
|
Moderately short | 0 | 1.9 | 11.1 | 24.4 | 12.5 | 12.5 | 2.4 | |
|
Post-winter growth habit |
Semi prostrate | 0 | 0 | 0 | 0 | 0 | 0 | 1.2 |
| Intermediate | 5.3 | 29.6 | 77.8 | 35.6 | 25.0 | 25.0 | 45.9 | |
| Semi erect | 75.4 | 50.9 | 22.2 | 31.1 | 50.0 | 62.5 | 37.6 | |
| Erect | 19.3 | 19.4 | 0.0 | 33.3 | 25.0 | 12.5 | 15.4 |
원산지 그룹별 주요 농업 형질들의 평균과 범위를 나타낸 것은 Table 3과 같다. 파종일에서 출수기까지의 일수를 나타낸 출수소요일수는 동아시아 유전자원 그룹이 195일로 출수가 가장 빨랐으며, 다른 모든 원산지 그룹과 통계적으로 유의미하게 구분되었다. 이는 한국을 비롯한 동아시아 지역에서는 트리티케일이 이모작으로 재배되기에 일찍 수확할 수 있는 조생종 특성이 요구되었기 때문인 것으로 생각된다. 초장 및 줄기 길이는 서유럽, 동아시아 원산지 그룹이 가장 긴 특성을 보인 반면, 북아메리카 원산지 그룹은 상대적으로 초장이 작았다. 이삭 길이는 원산지 그룹 간에 차이가 나타나지 않았다. 지엽 길이에서는 북·동유럽과 CIMMYT 원산지 자원들이 가장 길었고, 오세아니아·아프리카 원산지 그룹이 가장 짧았다. 천립중은 동아시아와 CIMMYT 원산지의 평균이 50 g으로 가장 무거웠고, 북·동유럽 원산지 그룹이 42 g으로 가장 가벼운 집단으로 분류되어 원산지 지역에 따라 종실 발달 정도에 큰 차이를 보였다. 한국 원산 자원이 대부분을 차지하는 동아시아 그룹 평균 초장은 133 ㎝로 타지역 자원에 비해 길었고, 천립중은 50 g으로서 타지역 자원보다 대립 경향을 보이고 있다. 이러한 동아시아 지역 그룹 자원들이 갖는 특성은 조사료 수량성 측면에서 우수한 것으로 판단되었다.
Table 3.
Basic statistics for quantitative traits in the 253 triticale genetic resources.
| Trait | East Asia |
North America |
Latin America (CIMMYT) |
Northern & Eastern Europe |
Western Europe |
Oceania & Africa | Total | |
| (n=57) | (n=108) | (n=27) | (n=45) | (n=8) | (n=8) | (n=253) | ||
|
Days to heading (days) | Meanz | 195d | 209b | 201c | 217a | 211ab | 216ab | 207 |
| Minimum | 186 | 194 | 194 | 195 | 205 | 206 | 186 | |
| Maximum | 215 | 236 | 211 | 230 | 227 | 245 | 245 | |
| CV(%) | 3.1 | 3.6 | 2.8 | 3.7 | 3.6 | 5.7 | 5.1 | |
|
Plant height (㎝) | Mean | 133a | 121b | 132ab | 124ab | 144a | 120ab | 126 |
| Minimum | 104 | 61 | 69 | 68 | 120 | 76 | 61 | |
| Maximum | 172 | 175 | 161 | 157 | 168 | 158 | 175 | |
| CV(%) | 11.2 | 18.1 | 15.8 | 18.0 | 12.2 | 23.1 | 16.9 | |
|
Culm length (㎝) | Mean | 121a | 110b | 120ab | 112ab | 132a | 109ab | 114 |
| Minimum | 94 | 50 | 60 | 61 | 109 | 65 | 50 | |
| Maximum | 161 | 164 | 150 | 147 | 158 | 146 | 164 | |
| CV(%) | 12.2 | 19.8 | 17.0 | 19.5 | 13.5 | 25.0 | 18.3 | |
|
Spike length (㎝) | Mean | 11.7a | 11.0a | 11.5a | 11.9a | 12.1a | 10.8a | 11.4a |
| Minimum | 8.9 | 6.2 | 8.5 | 7.0 | 9.8 | 7.0 | 6.2 | |
| Maximum | 13.8 | 18.0 | 14.3 | 17.3 | 17.0 | 12.6 | 18.0 | |
| CV(%) | 10.1 | 16.4 | 13.9 | 19.4 | 18.3 | 15.8 | 15.8 | |
|
Flag leaf length (㎝) | Mean | 16.1b | 16.5b | 18.9a | 19.6a | 17.9ab | 14.9b | 17.2 |
| Minimum | 12.5 | 9.8 | 13.5 | 12.1 | 12.7 | 6.8 | 6.8 | |
| Maximum | 20.3 | 24.0 | 25.2 | 27.3 | 25.0 | 20.3 | 27.3 | |
| CV(%) | 11.8 | 18.9 | 17.4 | 20.0 | 20.9 | 29.7 | 19.8 | |
|
Flag leaf width (㎝) | Mean | 1.4ab | 1.3bc | 1.4a | 1.3abc | 1.4abc | 1.2c | 1.3 |
| Minimum | 1.0 | 0.8 | 1.1 | 0.9 | 1.1 | 0.7 | 0.7 | |
| Maximum | 1.9 | 2.0 | 1.8 | 1.7 | 1.7 | 1.4 | 2.0 | |
| CV(%) | 13.0 | 14.3 | 14.5 | 14.2 | 14.1 | 18.9 | 14.5 | |
|
Thousand-grain weight (g) | Mean | 50a | 46b | 50ab | 42c | 46abc | 44abc | 47 |
| Minimum | 37 | 18 | 35 | 32 | 41 | 21 | 18 | |
| Maximum | 64 | 62 | 63 | 56 | 56 | 56 | 64 | |
| CV(%) | 12.5 | 19.1 | 11.2 | 14.9 | 11.5 | 25.4 | 17.1 |
상관분석
내한성과 주요 농업적 형질 간의 연관성을 파악하기 위해 상관관계를 분석한 결과는 Table 4와 같다. 내한성과 초장의 상관계수는 r = -0.17**, 내한성과 줄기길이 상관계수는 r = -0.18**로 음의 상관관계를 보였다. 본 내한성 조사에서 내한성 점수 척도가 낮으면 내한성이 강함을 의미한다. 따라서 내한성이 강할수록 초장이 길고 줄기가 긴 상관관계가 있음을 나타낸다고 할 수 있으며, 조사료용으로 적합한 줄기가 긴 자원들이 추위에도 잘 견딜 가능성이 있음을 시사한다. 내한성과 천립중은 상관관계가 거의 0에 가까워 내한성이 알곡의 무게와 상관이 없음을 나타냈다. 농업적 형질 간의 주요 상관관계를 살펴보면 출수소요일수와 천립중의 상관계수는 r = -0.60***로 중간 정도 음의 상관관계를 보였다. 출수기가 늦어질수록 천립중이 가벼워지는 경향이 뚜렷하였는데 이는 봄에 온도가 급격히 상승하고 건조한 기상 유형을 보이는 국내 기후 특성 때문에 생육 후기에 고온이나 수분 스트레스 등이 등숙에 영향을 주었을 가능성이 있으며, 조기 출수형 자원이 대립종 확보에 유리할 수 있음을 보여준다. 초장과 줄기길이 상관계수는 r = 1.00***으로 거의 완벽한 상관관계를 보이며, 이는 트리티케일의 초장 결정이 줄기길이가 주도하고 있음을 확인할 수 있었다. 지엽길이와 지엽폭은 양의 상관관계(r = 0.47***)를 보이며, 잎이 긴 자원이 폭도 넓은 경향이 있어 조사료 품질 면에서 유리한 특성을 가질 가능성이 높을 것으로 판단되었다. 이러한 농업 형질 간 상관관계를 살펴 볼 때 내한성이 강한 자원들이 조사료 수량과 직결되는 초장 및 줄기길이에서도 우수한 특성을 지니고 있을 가능성이 높을 것으로 판단되었다.
Table 4.
Correlation coefficients among 9 quantitative traits in 253 triticale genetic resources.
| Trait | DHy | PHx | CLw | SLv | ALu | FLt | FWs | TWr |
| WHz | -0.05 | -0.17** | -0.18** | 0.03 | -0.01 | 0.12 | -0.12 | 0.00 |
| DH | -0.15* | -0.16* | 0.08 | 0.08 | 0.24*** | -0.07 | -0.60*** | |
| PH | 1.00*** | 0.24*** | 0.01 | 0.20** | 0.19** | 0.18** | ||
| CL | 0.16* | 0.03 | 0.18** | 0.17** | 0.19** | |||
| SL | -0.12 | 0.28*** | 0.29*** | -0.08 | ||||
| AL | 0.05 | 0.03 | -0.10 | |||||
| FL | 0.47*** | -0.11 | ||||||
| FW | 0.20** | |||||||
| TW |
주성분분석
트리티케일 유전자원의 주요 농업 형질에 대한 주성분분석(PCA) 결과는 Table 5와 같다. 주성분 1의 설명력은 27.0%, 주성분 2의 설명력은 19.7%, 주성분 3의 설명력은 14.3%, 그리고 주성분 4의 설명력은 12.1%로 나타나 이 4개의 주성분으로 전체 변이의 73.2%를 설명할 수 있는 것으로 나타났다.
Table 5.
Eigenvalues and contributions obtained from principal component analysis in the 253 triticale genetic resources.
전체 변이의 73% 이상을 설명하는 제1 ∼ 제4주성분(PC1 ∼ PC4)과 주요 농업 형질 간의 상관관계를 분석한 결과는 Table 6과 같다. 제1주성분과 초장(r = -0.90***), 줄기길이(r = -0.89***)는 매우 높은 음의 상관을 보이기 때문에 제1주성분 값이 낮을수록 초장이 긴 자원임을 의미하기에 제1주성분을 초장을 설명할 수 있는 지표로 규정하였다. 제2주성분은 출수소요일수(r = 0.77***)와 강한 양의 상관, 천립중(r = -0.69***)과 강한 음의 상관을 보였다. 출수가 늦은 자원일수록 천립중이 가벼워지는 경향을 제2주성분이 설명해주고 있으므로 제2주성분은 출수기 및 종실 무게를 설명할 수 있는 지표로 규정하였다. 제3주성분은 지엽폭(r = 0.55***)과 내한성(r = 0.46***)에 높은 상관을 보여, 제3주성분은 잎 크기와 내한성을 설명할 수 있는 지표로 규정하였다. 제4주성분은 까락길이(r = 0.90***)와 매우 높은 상관을 보였다. 앞선 주성분들과는 독립적으로 나타나는 까락길이의 고유한 변이를 제4주성분이 설명하고 있어 제4주성분을 까락길이를 설명할 수 있는 지표로 규정하였다.
Table 6.
Four principal components among nine quantitative traits in the 253 triticale genetic resources.
| Traits | PC1 | PC2 | PC3 | PC4 |
| zWH (Score) | 0.21*** | 0.04 | 0.46*** | -0.21*** |
| yDH (Days) | 0.28*** | 0.77*** | -0.35*** | 0.18** |
| xPH (㎝) | -0.90*** | -0.04 | -0.37*** | -0.06 |
| wCL (㎝) | -0.89*** | -0.08 | -0.39*** | -0.03 |
| vSL (㎝) | -0.39*** | 0.46*** | 0.23*** | -0.36*** |
| uAL (㎝) | -0.03 | -0.03 | 0.00 | 0.90*** |
| tFL (㎝) | -0.39*** | 0.64*** | 0.37*** | 0.15* |
| sFW (㎝) | -0.52*** | 0.28*** | 0.55*** | 0.18** |
| rTW (g) | -0.37*** | -0.69*** | 0.43*** | 0.12 |
주성분분석에서 기여도가 가장 높은 두 개의 주성분 PC1을 가로축, PC2를 세로축으로 하여 트리티케일 유전자원 분포를 2차원 공간에 배열하여 시각화하였다(Fig. 1). 유전자원 변이의 27.0%를 설명하는 제1주성분인 가로축(x축)의 좌측으로 초장이 긴 자원들이 군집하고, 우측으로 까락길이가 긴 자원이 군집하였다. 전체 변이의 19.7%를 설명하는 제2주성분인 세로축(y축)에 따라 위 방향으로는 출수기가 늦은 유전자원들이 군집하고, 아래 방향으로는 출수기가 빠르고 천립중이 무거운 유전자원들이 군집하였다.
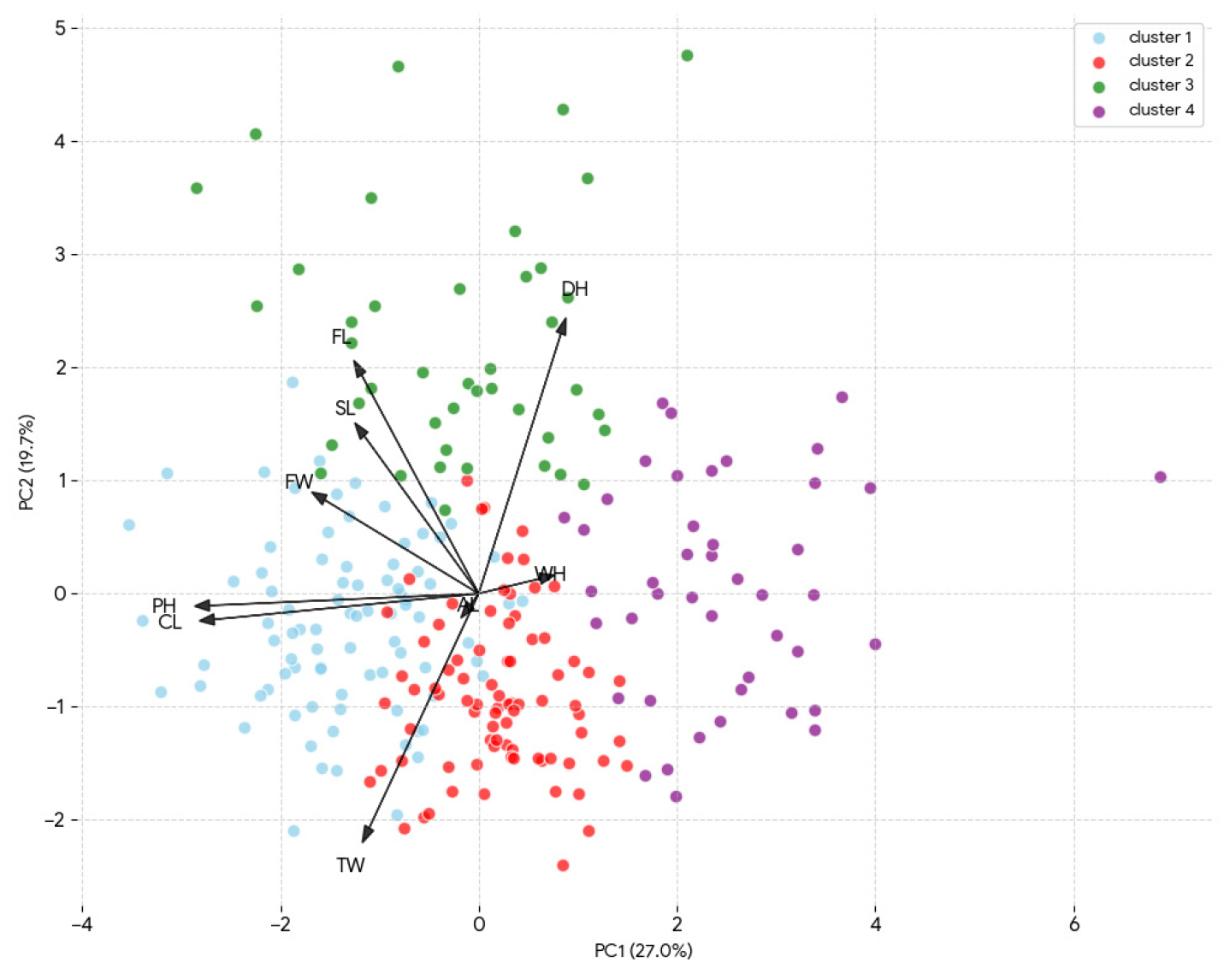
Fig. 1.
Principal component analysis (PCA) of agronomic traits and winter hardiness in the 253 triticale germplasms. A biplot showing variables and individuals in the first two principal components. WH, Winter Hardiness; DH, Days to Heading; PH, Plant Height; CL, Culm Length; SL, Spike Length; AL, Awn Length; FL, Flag leaf Length; FW, Flag leaf Width; TW, Thousand-grain Weight.
군집분석
트리티케일 자원들을 특성별로 그룹화하기 위하여 주성분분석으로 추출된 주성분 점수를 기반으로 Ward의 연결법과 유클리드 거리를 이용하여 유사한 자원들끼리 묶는 군집분석을 수행하였다(Fig. 2). 군집 Ⅰ은 전체의 19.0%에 해당하는 48개 자원이 속하였고, 군집 Ⅱ는 전체의 49.6%에 해당하는 125개 자원, 군집 Ⅲ은 전체의 14.3%에 해당하는 36개 자원, 그리고 군집 Ⅳ는 전체 유전자원의 17.1%를 차지하는 43개 자원이 속하였다. 4개 군집 간 양적 형질의 항목별 평균값의 결과는 Table 7과 같다. Ⅰ군집의 특징은 다른 군집보다 출수기가 가장 빨라 조기 수확에 유리하며, 천립중이 50 g으로 가장 무거워 종실용으로 적합한 특성을 보였으나 내한성이 가장 약한 유전자원이 속한 군집이었다. Ⅱ군집은 평균 초장이 136 ㎝로 가장 길어 조사료 수량 확보에 유리한 형질을 지녔고, 내한성이 다른 그룹보다 가장 강하였으며, 천립중도 49 g으로 Ⅰ군집과 비슷한 무게를 갖는 곡실 특성을 보유한 군집이었다. Ⅲ군집은 초장이 90 ㎝ 정도로 작고 출수기가 늦어 조사료 생산성 측면에서는 불리한 형질을 지녔으나 종실 생산 측면에서는 도복저항성 특성을 도입할 수 있는 자원이 속하였다. Ⅳ군집은 출수소요일수가 219일로 출수기가 가장 늦으며, 지엽의 길이는 20.8 ㎝로 가장 길었다. 또한 이 군집에서는 잎의 면적이 커서 풍엽성 특성을 나타내어 조사료의 품질(가소화영양소, 조단백함량 등)을 높일 수 있는 잠재력이 높은 자원들이 포함되어 있었다.
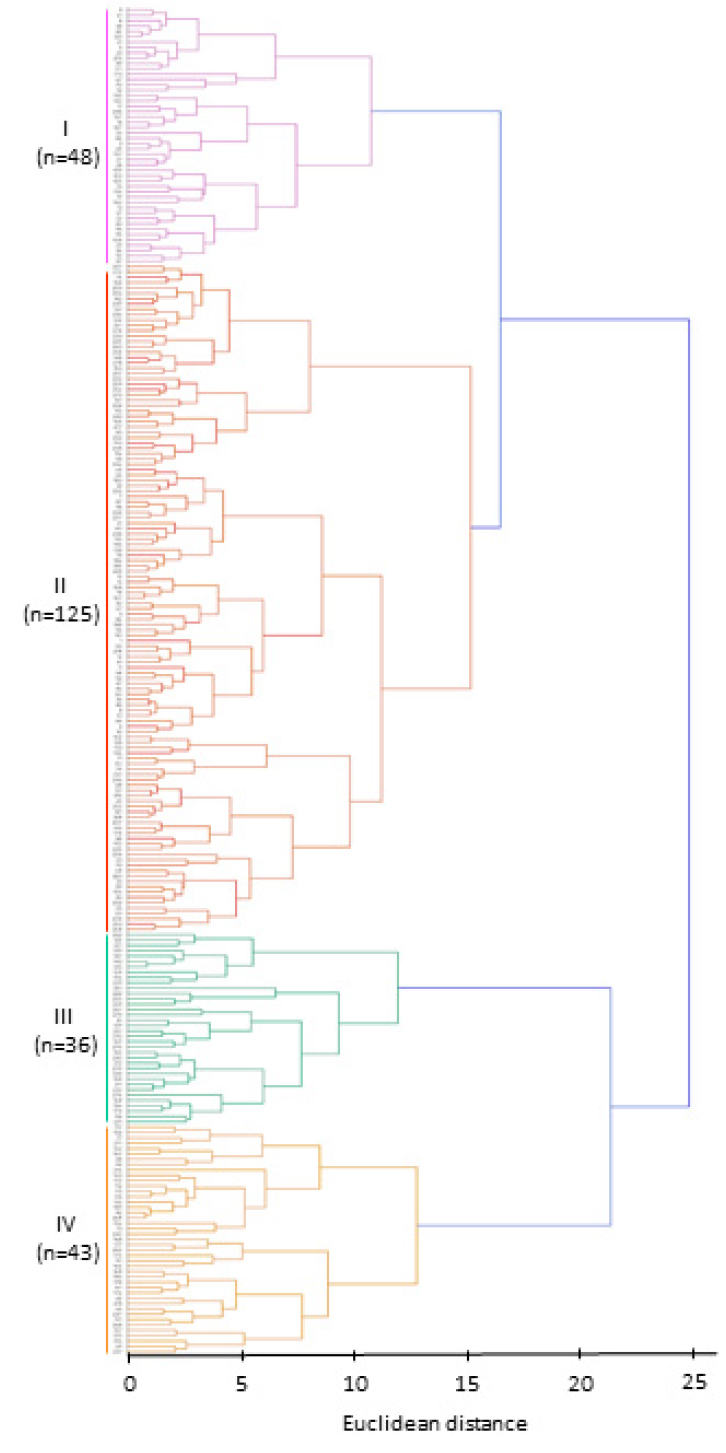
Table 7.
Statistical mean values of winter hardiness and agronomic traits for each cluster based on Ward’s method.
| Characters | Cluster (mean) | |||
| I | II | III | IV | |
| (n=48) | (n=125) | (n=36) | (n=43) | |
| Winter hardiness (1-9 score)z | 6 | 3 | 4 | 5 |
| Days to heading (Days) | 198 | 204 | 213 | 219 |
| Culm length (㎝) | 112 | 125 | 79 | 116 |
| Spike length (㎝) | 11.5 | 11.2 | 10.4 | 12.8 |
| Awn length (㎝) | 4.4 | 5.3 | 5.4 | 6.2 |
| Plant height (㎝) | 124 | 136 | 90 | 129 |
| Flag leaf length (㎝) | 17.8 | 16.2 | 15.7 | 20.8 |
| Flag leaf width (㎝) | 1.4 | 1.3 | 1.3 | 1.4 |
| Thousand-grain weight (g) | 50 | 49 | 42 | 40 |
고 찰
트리티케일은 밀과 호밀의 합성 작물로, 복잡한 유전적 배경을 가지는 경우가 많기 때문에 수집된 유전자원의 보존, 특성 평가 및 분류를 통해 유용한 특성 발굴, 유전자원의 관리와 보존의 효율성 증대, 육종 전략 수립을 이룰 수 있다(Furman et al., 1997). 우리나라에서 재배되는 트리티케일과 벼 작부체계는 벼 수확 후 10월에 파종하고 이듬해 벼 이앙 전인 5월에 조사료용으로 수확하는 이모작 방식이다. 최근 국내 기후변화 변동이 잦아지고 있는 추세이며(Park and Suh, 2023), 겨울철 기온은 상승하고 있으나 한파 발생 빈도는 증가하고 있다(Jeon and Cho, 2015). 따라서 국내 트리티케일 품종개발에서 요구되는 육종 목표는 출수기까지의 소요일수 단축과 더불어 내한성을 강화하는 것이다.
전체 유전자원에서 내한성 등급 분포에서 내한성 ‘매우 강함’과 ‘강함’ 등급 비율이 과반을 차지하고 있다(Table 2). 특히 북미, 동아시아, 북·동유럽에 내한성이 강한 자원 비율이 높다는 점은 이 지역들이 겨울 추위가 심한 지역으로서 동해·저온 스트레스하에서 생존력이 높은 자원이 상대적으로 많이 포함되었다. 이러한 결과는 내한성 등급 비율이 유전자원의 기원군 지역에 따라 뚜렷이 달라질 수 있으며, 특히 한랭지 기원군에서 내한성 강한 자원의 축적이 확인된다는 Prášil et al. (2025)의 보고와 같았다. 잎색을 비롯한 조직 기관의 착색 정도, 기관 형태, 월동 전 생장도와 월동 후 생장형 등의 질적 특성이 원산지 그룹 별로 분포 비율이 차이가 있었다. 이러한 결과는 유전자원의 집단 구조는 기원지의 기후와 육종 역사에 따른 적응 결과가 반영된다는 Badu-Apraku (2021)의 보고와 비슷한 유형이다. 이삭길이를 제외한 출수기, 초장, 줄기길이, 지엽길이와 폭, 천립중 등의 양적형질들에서 원산지 그룹 간 평균에서 통계적 차이가 인정되고 있다(Table 3). 이는 갈대에서 원산지의 위도와 재배환경 차이에 따른 형질 평균과 변이의 동시 변화는 지역에 대한 적응과 더불어 환경에 따른 유연한 형질 변화가 같이 작용할 수 있다(Ren et al., 2020)는 사례에서 보듯이 트리티케일 자원 기원지의 재배 시기, 온도, 수분 조건 등 재배환경과 육종 선택의 누적 결과에 기인한 것으로 생각된다.
주요 농업 형질들 간의 상관계수 표(Table 4)에서 출수소요일수와 천립중은 고도로 유의한 음의 상관관계(r = -0.60***)가 나타났다. 출수가 늦어질수록 천립중이 감소하는 생육시기와 곡립충실도 간의 상충관계(트레이드오프)가 본 집단에서 뚜렷하게 관찰되었다. Kim et al. (2024)에 의하면 국내에서 밀의 경우 지난 10년간 출수기는 매년 약 0.9일 앞당겨지고 천립중은 매년 0.2062 g 증가하는 경향이었다. 이는 고온, 가뭄 등 환경 스트레스가 등숙기에 겹칠 때 만숙 계통의 종실 충실이 불리해질 수 있음을 나타낸다. 초장과 줄기길이는 매우 강한 양의 공선성(r = 1.00***)을 보이고 있다. 이는 초장이 줄기길이에 의해 결정됨을 의미한다. 본 유전자원 집단에서는 내한성이 강한 자원일수록 초장이 큰 경향(r = -0.17***)이 나타나 내한성과 생체량이 부분적으로 연동된 패턴을 보였다. 내한성과 천립중과는 무상관(r = 0.00)으로 나타나 내한성과 천립중은 분리 선발할 수 있는 여지가 있음을 보였다. 다만 내한성은 다수의 유전자가 관여하고 상호작용 중 상위성(epistasis) 효과가 커서 단일 환경, 단년도 상관만으로 고정적 관계로 일반화하기 어렵다(Liu et al., 2014)는 보고가 있기에 내한성 도입 육종은 다년간에 걸쳐 선발이 이루어져야 할 것으로 생각한다.
농업 형질들의 정보압축을 위해 주성분분석을 한 결과 PC1과 PC2는 각각 27.0%와 19.7%의 변이를 설명하여 누적 46.7%였으며, PC4까지 누적 설명분산은 73.2%로 나타났다(Table 5). 이는 평가된 농업 형질들이 상호 독립적이라기보다 소수의 잠재 변이 축으로 요약될 수 있음을 의미한다(Bao et al, 2023; Naghavi and Jahansouz, 2005). 고유값이 1 이상인 4개의 주성분과 유전자원의 특성 간 상관관계(Table 7)를 분석한 결과 제1주성분은 초장(r = -0.90***), 줄기길이(r= -0.89***), 지엽폭(r= -0.52***)이 같은 방향인 음의 상관관계를 맺고 있어, 제1주성분은 전반적인 식물체 크기 변이 축으로 요약할 수 있다. Kroupin et al. (2019)에 의하면 춘파형 트리티케일에서 호밀에서 유래된 단간유전자(Ddw1) 도입은 초장 감소와 함께 출수 지연, 천립중 및 이삭이 작아지는 변화가 동반되어 초장 형질만 분리 선발이 어렵다고 보고하고 있다. 이는 도복저항성을 높이기 위하여 단간유전자를 도입할 경우 식물체 크기가 작아지는 방향으로 옮겨질 수 있어 식물체 크기가 커야 하는 조사료용 목적 육종에서는 단간유전자 도입은 배제해야 함을 의미한다. 제2주성분은 출수소요일수(r = 0.77***), 지엽길이(r = 0.64***), 이삭길이(r = 0.46***) 등이 양의 상관관계이고, 천립중(r = -0.69***)이 음의 상관관계를 이루고 있어 생육기간, 잎, 이삭 크기 병행 증가와 천립중(종실 비대) 감소의 대립관계 축을 형성하고 있다. 제3주성분은 내한성(r = 0.46***), 엽폭(r= 0.55***), 천립중(r= 0.43***)이 같은 방향으로 움직이는 축으로 요약된다. 내한성 척도 점수가 낮을수록 내한성이 강함을 나타내므로 내한성이 강한 방향으로의 선발은 잎이 좁아지고 천립중이 가벼운 경향으로 움직이는 것으로 해석할 수 있다. 내한성은 제3주성분에, 초장은 제1주성분에 크게 기여하는 형질로 나타났다. 서로 다른 주성분 간에는 독립성이 유지되므로, 이는 두 형질 간의 상관관계가 낮아 내한성과 초장을 각각 독립적으로 육종할 수 있음을 시사한다. 제4주성분은 다른 형질들과 비교하여 까락길이와 압도적으로 가장 큰 상관관계를 갖고 있어 다른 주성분들과 상대적으로 독립적인 변이 축을 형성하고 있다. 이 결과 또한 초장이 길고 가축기호성을 개선시킬 수 있는 까락이 없거나 짧은 까락을 지닌 품종을 개발할 수 있는 가능성이 있음을 나타낸다. 트리티케일 초장을 조절하는 QTL이 5A, 4B 염색체와 5R 염색체에 있고(Trini et al., 2021), 트리티케일 까락길이를 조절하는 QTL 위치는 아직 보고되지 않았지만 밀에서 까락길이 조절 유전자가 5A 염색체의 긴 팔 끝부분에 위치해 있다(Sun et al., 2024)는 보고는 초장 관여 유전자와 까락길이 관여 유전자가 멀리 떨어진 다른 위치에 존재하여 초장과 까락에 대해 독립적인 육종이 가능함을 뒷받침한다.
본 트리티케일 유전자원 집단을 대상으로 다변량 형질공분산을 바탕으로 Ward’s 계층적 군집을 한 결과 전체 집단을 4개의 군집으로 분류할 수 있다(Fig. 1, Fig. 2, Table 7). 군집 I (Cluster I)은 강한 내한성, 조숙성, 무거운 천립중을 동시에 갖춘 그룹으로서 국내 이모작 환경에 적합한 가장 직접적인 육종 소재가 될 수 있는 유전자원 집단이 될 수 있다. 군집 Ⅱ (Cluster Ⅱ)는 가장 긴 초형을 갖는 생체량 다수확 잠재력을 갖고 있지만 내한성은 다른 군집에 비해 가장 약한 유전자원 집단이다. 이 군집은 생체량 다수확 잠재력을 제공하는 공여친(parental donor)으로 유용할 것으로 생각되지만 이 군집에 속한 유전자원을 이용한 교배 후대에서는 내한성을 선발지수에 반드시 포함시키는 육종전략(Michel et al., 2019)이 필요할 것으로 생각된다. 군집 Ⅲ (Cluster Ⅲ)은 곡실생산 안정성을 높이기 위해 단간유전자를 도입하여 초장을 줄여 도복저항성을 높인 유전자원 그룹이 구성된 것으로 생각된다. 군집 Ⅲ에 포함된 단간형질 자원을 이용해 도복저항성을 증진하려면, 출수지연과 천립중 감소를 보상할 수 있도록 조숙·대립 계통과의 교배설계가 필수적인 것으로 보고된 바 있다(Kroupin et al., 2025). 군집 Ⅳ (Cluster Ⅳ)는 출수가 가장 늦고 까락 관련 값이 비교적 큰 반면 천립중이 가장 낮은 유전자원 집단이다. 가축기호성 개선을 위해 까락이 없거나 극히 짧은 품종을 개발하는 데 이 군집을 공여친으로 이용할 경우, 국내 재배환경에 맞도록 출수기를 앞당기는 선발이 필요하다. 주성분분석 결과 제1주성분부터 제4주성분까지가 전체 변이의 73.2%를 설명하므로, 단일 형질 선발보다는 체형 축과 출수-천립중 축 같은 형질 패키지 단위(Michel et al., 2019; Wooten et al., 2009)로 육종 소재를 선택하는 것이 합리적이다.
결과적으로 벼 - 트리티케일 이모작 작부체계에 적합한 우수 품종(출수가 빠르고 조사료 수량이 많으며, 내한성이 강한 품종)을 육성하기 위해서는 개화소요일수가 198일로 가장 짧은 Ⅰ군집의 자원과 내한성이 가장 강하고 초장이 125 ㎝로 가장 긴 Ⅱ군집에 속한 자원들을 교배 모부본으로 활용하는 것이 우리나라 농업환경에 적합한 트리티케일 품종개발에 가장 유용할 것으로 판단된다.
적 요
본 연구는 국립식량과학원에서 보유한 17개국 유래 253점의 트리티케일 유전자원을 대상으로 내한성 및 주요 농업적 형질의 다양성을 평가하여 국내 논 이모작 재배에 적합한 품종 육성을 위한 기초 자료를 확보하고자 수행되었다. 유전자원의 원산지에 따른 특성 분석 결과, 한국을 포함한 동아시아 그룹은 출수소요일수가 195일로 가장 짧아 조생종 특성이 뚜렷했으며, 평균 초장이 133 ㎝로 길고 천립중이 50 g으로 무거워 조사료 수량성 측면에서 우수하였다. 내한성 평가에서는 서유럽 지역 유전자원의 87.5%가 내한성이 강하였으며, 초장과 줄기길이가 긴 경향을 보여 추위에 강하면서도 조사료 생산성이 높은 자원 선발의 가능성을 확인하였다. 형질 간 상관관계에서는 출수기가 늦어질수록 천립중이 가벼워지는 경향(r = -0.60***)이 뚜렷하게 나타나 국내 기후 환경에서 조기 출수형 자원이 대립종 확보에 유리함을 나타냈다. 주성분분석을 통해 전체 변이의 73.2%를 설명하는 4개의 주성분을 도출하였으며, 제1주성분은 초장 및 줄기길이와, 제2주성분은 출수기 및 종실무게와 밀접한 연관성을 보였다. 군집분석 결과, 출수기가 가장 빨라 조기 수확에 유리한 군집 Ⅰ, 내한성이 가장 강하고 초장이 길어 조사료 생산성이 우수한 군집 Ⅱ, 초장이 작고 출수기가 늦은 군집 Ⅲ, 그리고 지엽이 길고 까락이 짧아 조사료 품질 향상에 유리한 특성을 지닌 군집 Ⅳ로 분류되었다. 따라서 국내 논 이모작 작부체계에 적응하고 내한성 다수확 조사료 특성을 지닌 우수 트리티케일 품종을 육종하기 위해서는 출수기가 빠른 군집 Ⅰ의 자원과 내한성이 강하고 초장이 긴 군집 Ⅱ의 자원 위주로 교배 모부본으로 활용하는 것이 가장 유용할 것으로 판단되었다.
