

서 언
재료 및 방법
실험재료의 재배관리, 재배지대 및 유형
기상자료 수집
개화단계 설정 및 개화 특성 조사
생육특성 조사 및 정유 추출
통계분석
결과 및 고찰
개화단계와 형태학적 특성
개화시기와 기상과의 관계
재배지대와 유형별 생육량의 변화
재배지대와 유형별 개화 및 생육량 비교
적 요
서 언
꿀풀과(Lamiaceae) 향유속(Elsholtzia)에는 최소 33종 이상이 있으며, 아프리카, 북미, 유럽, 아시아 국가 등에 널리 분포되어 있다(Guo et al., 2012). 국내에는 외부 형태학적 분류 기준으로 가는잎향유(E. angustifolia), 향유(E. ciliate), 좀향유(E. minima), 꽃향유(E. splendens), 한라꽃향유(E. hallasanensis), 애기향유(E. saxatilis) 6종이 서식하는 것으로 알려져 있다(Lee, 2000).
이 중 꽃향유는 주로 방향성 식물로 민간요법, 허브 티(herb tea), 식품, 향신료, 음료, 향수, 화장품, 아로마테라피의 원료로 사용되어 왔다(Guo et al., 2012). 특히 중국에서는 꽃향유가 구리 내성 식물로써 토양 호흡 증가와 효소 활성 강화를 통해 구리를 체내에 저장하는 특성(Yang et al., 2015)을 가지고 있어 중금속에 오염된 토양에서 초기 우점(dominance) 식물로 이용되기도 한다(Zhang et al., 2010a, b).
최근 치유농업 관련 법률이 시행되면서 각기 다른 소비계층을 대상으로 심신 안정화를 목적으로 한 농업 연계 활동이 상업화되고 있으며, 관련 프로그램 개발을 위해 활용도가 높은 국내 자생식물에 대한 관심이 커지고 있는 상황이다. 꽃향유는 소염, 진통, 해열, 항균 효과(Yoon, 1992)가 있어 국내에서 한약재로 사용되어 왔으며(Kim et al., 1993), 정유에는 항산화 성분과 naginata ketone, linalool, limonene 등의 방향성 물질이 함유되어 있어 천연 향료로써 개발 가능성이 높다. 또한 자줏빛의 수상화서가 군집을 이루어 있어 관상용(Jeon and Hong, 2006)은 물론, 중금속에 의해 훼손된 토양의 식생 회복을 위해서도 활용이 가능하다(Ye et al., 2002).
반면 꽃향유와 관련하여 일부 개화유도 관련 보고(Chang et al., 2003; Sohn and Kim, 2003) 외에는 효능과 함유 성분에 관한 연구가 대부분이며, 주 분포지대가 해발고도 200~300 m 이상의 고지대인 것으로 알려져 있으나(Wu et al., 1977) 다양한 환경에서 재배 적응성에 관한 연구가 전무한 실정이다. 따라서 본 연구는 꽃향유의 재배지대와 재배유형에 따른 개화 및 생육 특성에 대한 기본 재배정보를 조사·분석함으로써 재배적응성을 평가하여 농산업적 활용도 확대를 위해 수행되었다.
재료 및 방법
실험재료의 재배관리, 재배지대 및 유형
꽃향유 종자는 2019년 전북특별자치도 임실군에서 농업인이 채취하여 보관중인 종자를 분양 받아 2022년 연구소 내에서 자가 채종한 종자를 이용하였다. 실험에 사용할 묘 생산을 위해 1공의 용적이 30 mL 인 105공 plug tray에 원예용상토(등록번호: 전북 김제014-가-11014)를 이용하여 2023년 4월 17일에 파종하였고, 평균 초장 15.5±1.5 ㎝, 주경 1.27±0.11 ㎜, 본엽 6매가 전개된 건전하고 균일한 묘를 정식에 사용하였다. 육묘관리를 위해 전북특별자치도농업기술원 허브산채시험장 육묘 온실의 평균온도를 25~30℃ 범위로 조절하였다.
본 시험포장으로의 정식은 모든 처리구(재배지대, 유형)에서 5월 18일 동일한 날 50×50 ㎝ 간격으로 정식하였으며, 정식 전 시험포의 기비는 작물별 비료사용처방(RDA, 2019)의 1년생 초화류에 준하여 처리하였다. 토양수분 관리는 점적관수 형태로 모든 시험장소에 텐시오미터(Model SR, Irrometer Co., USA)를 설치하여 –45 kPa 도달 시 관수하였다.
재배지대 중 평야지는 전라북도 익산시 신흥동(35°56’27”N, 126°59’27”E, 해발고도 12 m), 중산간지는 남원시 운봉읍(35°24’44”N, 127°31’36”E, 해발고도 507 m)에 위치한 농업기술원 시험포장을 활용하였으며, 재배유형은 지대별 각각 노지와 비가림하우스에 정식한 후 개화 및 생육 특성에 대한 조사를 진행하였다.
기상자료 수집
재배지대와 유형별로 solar radiation shield를 부착한 실외용 온·습도 데이터로거(U23-001, Onset Computer Corp., USA)와 온도 데이터로거(UA-001-08, Onset Computer Corp., USA) 2개를 지면으로부터 약 1.5 m 높이에 각각 설치하여 정식일로부터 수확일까지의 온도 데이터를 1시간 단위로 수집하였다. 수집한 온도데이터는 일평균온도와 함께 기준온도(reference temperature)를 10℃로 설정한 후 누적하여 개화기까지의 누적 적산온도(effective accumulated temperature)를 산출하였다.
또한 일조시간(duration of sunshine)은 기상청(KMA, Korea Meteorological Administration)의 자동기상관측시스템(automatic weather system)이 설치된 관측지점 146 (전주), 247 (남원)으로부터 수집하였으며, 정식일부터 누적하여 누적 일조시간(integrated sunshine hours)을 산출하였다.
개화단계 설정 및 개화 특성 조사
개화단계는 화뢰가 관찰되는 화뢰출현기(flower bud emergence), 수상화서(spike)가 신장하는 화뢰신장기(flower bud elongation), 화서의 40% 이상이 개화한 개화기(flowering period), 70% 개화한 만개기(full bloom), 30%의 낙화가 관찰되는 낙화기(late bloom)로 구분하였다. 또한 개화단계 중 수확기는 낙화기를 기준으로 결정하였는데, 개화기에 화수의 생성과 함께 정유 함량이 높아진다는 보고(Putievsky et al., 1986; Werker et al., 1985)와 생육단계 중 낙화기에 정유 추출량이 많았던 사전 실험 결과(데이터 미제시)를 근거로 설정하였다.
수상화서의 길이는 꽃이 착생한 화서의 하단부터 상단부 끝까지의 길이로 측정하였고, 폭은 길이의 1/2 지점을 2회 측정하여 평균으로 산출하였으며, 화서경의 두께도 동일한 방법으로 조사하였다. 주당 화서수는 생육이 균일한 10개체를 대상으로 조사하였고, 화서당 소화의 개수는 임의로 채취한 화서 30개를 대상으로 착생한 모든 소화를 계수하였다.
생육특성 조사 및 정유 추출
재배지대와 유형별 정식일수에 따른 생육조사 중 초장은 지제부에서 주경(main stem) 정단부까지를 측정하였고, 주경은 지면 위 5 ㎝ 높이의 줄기 직경을 2회 측정하여 평균으로 산출하였다. 분지수는 주경으로부터 발생한 1차 분지수와, 1차 분지로부터 발생한 2차 분지수를 계수한 후 합산하여 표시하였다. 조사기간 내 동일한 10개체를 대상으로 조사하였으며, 정식 후 0~138일까지 10일 간격으로 15회 조사하였다.
수확기 초장, 초폭, 주경, 분지수는 정식일수별 생육조사와 동일한 방법으로 조사하였다. 수확한 식물체 전초는 개체별로 생체중을 측정하였고, 음건한 후 건물률은 생체중을 건물중으로 나눈 값을 백분율로 산출하였다. 또한 정유(essential oil) 추출은 각 재배지대와 유형별 개화종 단계에서 수확하여 건조(음건)한 후 분쇄된 시료 500 g을 대상으로 SDE (simultaneous distillation extraction)법(RDA, 2013)을 적용하여 10반복으로 추출량을 측정하였다.
통계분석
데이터의 분석과 그래프 작성은 통계프로그램(Sigmaplot 14.0, Systat Inc., USA)을 활용하였다. 개화단계는 1월 1일 기준 Julian day로 환산하여 평균간 다중비교(DMRT)하였고, 생육시기별 초장, 경경, 분지수는 산술평균과 표준오차를 산출하였다. 또한 수확기 개화, 생육 및 수량특성 항목을 대상으로 산술평균, 표준오차 산출 및 요인분석을 실시하여 유의성을 검정하였다.
결과 및 고찰
개화단계와 형태학적 특성
꽃향유를 2023년 5월 18일 정식하여 재배지대(평야지, 중산간지)와 유형(노지, 비가림하우스)별 개화단계를 조사한 결과(Table 1), 중산간지에서 평야지보다 화뢰 출현(Fig. 1A)이 2~3일, 개화기(Fig. 1C) 9일, 만개기(Fig. 1D)는 6~7일 빨랐다. 재배유형에 따라서는 평야지 노지와 비가림하우스에서 각각 10월 9일과 10월 11일로 노지재배에서 약 2일 빠른 경향을 보였으며, 이와 같은 결과는 중산간지 재배유형과 화뢰 출현, 만개기에서도 동일한 경향을 보였다. 화뢰 출현부터 개화까지의 소요일수는 중산간지 노지재배에서 27일로 가장 짧았으며, 중산간지 비가림하우스(31일), 평야지 노지재배(34일), 비가림하우스(37일) 순으로 평균 약 32일 소요되었다.
Table 1.
The effects of different altitudes and cultivation methods on flowering stage of Elsholtzia splendens with the same planting date in 2023.
|
Different altitudes |
Cultivation methods |
Planting date |
Flower bud emergence (Julian day) |
Flowering period (Julian day) |
Full bloom (Julian day) |
Late bloom stage (Julian day) |
| Plains | Field | May 18 | Sep. 5 (247 az) | Oct. 9 (281 b) | Oct. 15 (287 a) | Oct. 22 (294 b) |
| Plastic house | May 18 | Sep. 4 (246 ab) | Oct. 11 (283 a) | Oct. 16 (288 a) | Oct. 24 (296 a) | |
| Mid-mountain | Field | May 18 | Sep. 3 (245 b) | Sep. 30 (272 d) | Oct. 8 (280 c) | Oct. 16 (288 d) |
| Plastic house | May 18 | Sep. 1 (243 c) | Oct. 2 (274 c) | Oct. 10 (282 b) | Oct. 18 (290 c) |
꽃향유의 꽃의 특성은 많은 소화가 한쪽으로 치우쳐 착생한 2~10 ㎝ 길이의 수상화서가 원줄기 또는 측지 끝에 착생하고, 홍자색의 꽃을 피우는 특성(Jeon and Hong, 2006)을 보였다(Fig. 1F). 또한 꽃받침은 종형 또는 원추형의 형태로 4개의 수술 중 보통 2~3개가 길며(Fig. 1G), 주두는 화관 밖으로 돌출되어 있는 형태를 보이는데(Li and Hedge, 1994; Press, 1982), 본 연구에서 관찰된 화서와 소화의 모습도 유사한 결과를 보였다.
식물체의 전체 형태는 원줄기에서 1차 측지의 분지, 1차 측지에서 2차 측지가 연속적으로 분지하여 전체적으로 원뿔형을 뒤집어 놓은 듯한 형태를 보였으며(Fig. 1I), 최대 4차까지의 분지 발생 후 일시에 화뢰가 출현, 개화하는 특성을 나타내었다. 또한 식물체 수관 내 하단부의 주경과 측지의 기부에서 부정근 발생(Fig. 1H)이 많았는데, 이는 대체로 노지보다 비가림하우스에서 많은 편이었다.

개화시기와 기상과의 관계
재배지대와 유형별 정식기부터 수확기까지의 평균온도(Fig. 2A)는 평야지 비가림하우스 25.7℃, 노지 24.1℃, 중산간지 비가림하우스 22.6℃, 노지 21.2℃로 조사되었다. 특히 유효적산온도(Fig. 2B)는 평균온도에 따라 각각 2,423.2℃, 2,149.7℃, 2,009.1℃, 1,779.4℃, 평균 2,090.4℃로 조사되었다. 그러나 유효적산온도가 낮은 중산간지 노지 재배에서 개화기가 빨랐는데(Table 1), 잇꽃(Choi et al., 2024)의 재배고도가 높을수록 개화기가 늦어진다는 보고와 반대의 결과를 나타내어 온도는 개화시기와의 유의한 관계는 없는 것으로 추정되었다. 반면 재배지대와 유형에 따른 개화기 차이(6~7일)는 화뢰 출현기의 차이(2~3일)보다 컸는데, 이는 국화에서 화뢰 출현부터 개화까지의 소요일수가 적온 이상의 고온 조건에서 지연된다는 보고(Nagasuga et al., 2008; 2013)와 유사한 경우로 생각되었다.
평균 일조시간(Fig. 2C)은 정식일인 5월 18일부터 6월 19일까지 평야지와 중산간지에서 각각 7.6, 7.0시간으로 조사되었으나, 장마가 시작된 6월 20일부터 화뢰 출현기까지는 각각 5.6, 5.1시간으로 짧아졌다. 정식일부터 화뢰 출현기까지의 누적 일조시간은 평야지 노지와 비가림하우스에서 각각 690.2, 682.2시간, 중산간지 노지와 비가림하우스에서 각각 617.0, 608.7시간으로 조사되어 온도에 비해 재배지대와 유형별 개화시기가 유사한 특성을 나타내었다. 또한 추가적인 일조시간 분석을 통해 6월 20일부터 화뢰 출현까지 소요된 73~77일 중, 일조시간이 8시간 미만인 일수는 평야지 노지와 비가림하우스 재배에서 50일, 중산간지 노지와 비가림하우스 재배가 각각 59, 57일로 조사되었다.
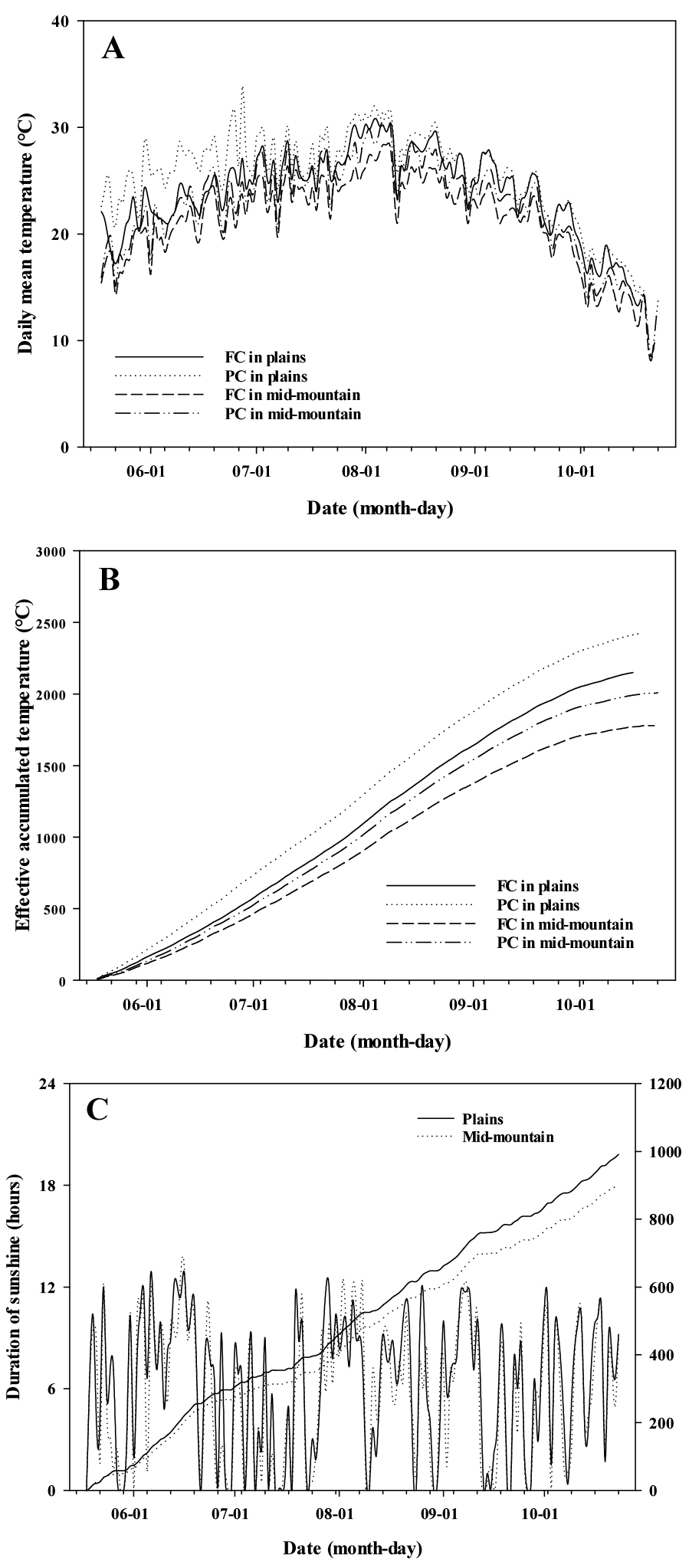
Fig. 2.
The changes in daily mean temperature (A), effective accumulated temperature (B), duration of sunshine, and integrated sunshine hours (C) according to the different altitudes and cultivation methods at experimental regions where Elsholtzia splendens are grown. FC and PC in the legend represent field cultivation and plastic house cultivation, respectively.
꽃향유의 개화는 춘화 조건 없이 단일 광주기에 의해 유도되며(Chang et al., 2003), 본엽이 최소 4~6매 이상 전개된 상태에서 8~9시간의 단일처리 약 30~34일 후 개화가 시작된다(Sohn and Kim, 2003). 본 연구 결과에서도 개화반응과 온도와의 연관성은 보이지 않았으나 중산간부에서 평야부보다 화뢰 출현과 개화가 빨랐던 것은 온도보다 일조시간(일장)에 더 반응했을 것으로 추정되었다. 또한 장마기부터 시작된 일조시간의 감소는 평야지와 중산간지 에서 50~59일 경과되어 선행연구 결과와 차이를 보였다. 이는 화분 실험을 통해 연속적 단일처리를 한 선행 연구와 달리 본 연구에서는 자연적 환경하에서 달라진 일조시간이 개화를 유도하는 단일조건에 영향을 주었을 것으로 생각되었다.
재배지대와 유형별 생육량의 변화
재배지대와 유형에 따라 꽃향유의 정식 후 일수별 생육 조사 결과(Fig. 3), 초장(Fig. 3A)은 정식 후 19일까지는 큰 차이가 없었으나 정식 후 29일경부터 처리간 차이를 보이기 시작하였다. 정식 138일 후 평야지와 중산간지 비가림하우스에서 초장은 143.2 ㎝로 생육량이 많았으며, 다음으로 평야지 노지(94.7 ㎝), 중산간지 노지(80.1 ㎝) 순이었다. 주경의 경우 신초장과 유사한 생육 양상을 보였으며, 정식 후 138일에는 평야지 비가림하우스(22.4 ㎜), 중산간지 비가림하우스(18.4 ㎜), 평야지 노지(14.3 ㎜), 중산간지 노지(9.0 ㎜) 순으로 신초장보다 처리간 차이가 뚜렷하였다(Fig. 3B).
1차 분지 수(Fig. 3C)는 정식 후 59~69일경까지, 1, 2차 분지 수의 합(Fig. 3D)은 정식 후 29일부터 급격히 증가하기 시작하여 정식 후 59일부터는 완만한 경향을 보였으며, 처리간 분지 수의 차이는 주경과 동일한 결과를 나타내었다. 재배지대와 유형에 따른 영양생장량의 차이는 생육온도(Fig. 2A)에 기인한 것으로 생각되며, 12~28℃의 범위에서 생육온도가 높을수록 초창과 초폭 등 기본 영양생장량이 많았다는 Chang et al. (2003)의 보고와 유사하였다.
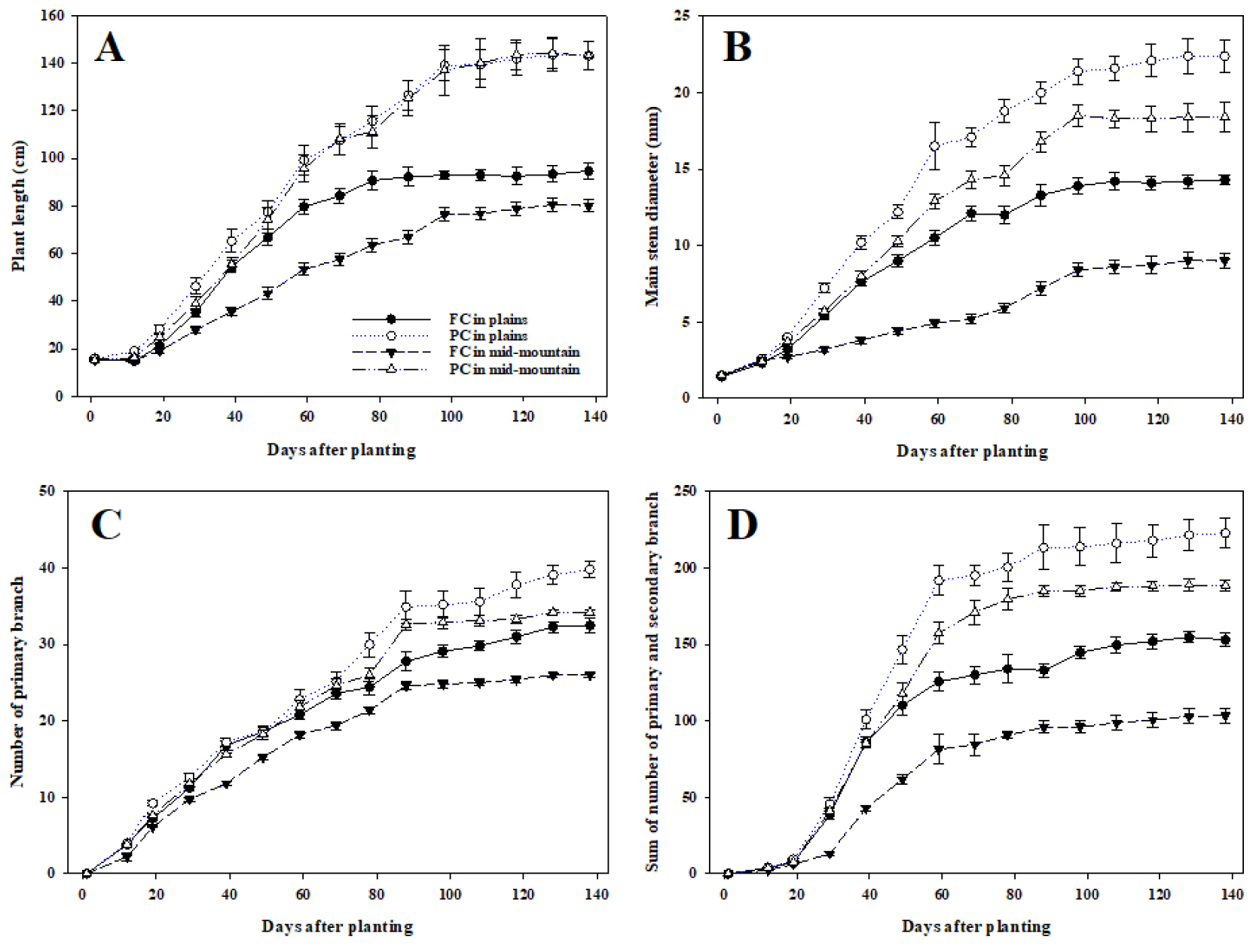
Fig. 3.
The changes in plant length (A), main stem diameter (B), number of primary branches (C), and sum of number of primary and secondary branches (D) according to the different altitudes and cultivation methods of Elsholtzia splendens. Vertical bars represent standard error of the means (n = 10). FC and PC in the legend represent field cultivation and plastic house cultivation, respectively.
영양생장에서 생식생장으로의 전환은 현화식물에서 가장 중요한 발달 단계이다(Wang et al., 2020). 일반적으로 여러 초본 화훼류, 과채류, 낙엽 과수 등에서 영양생장과 생식생장 기관간에는 생장상관 관계를 나타내며, 결과적으로 영양생장이 종료되는 시점에 생식생장으로의 발육이 진행된다(Pyon et al., 2014). 본 연구에서도 정식 후 59~69일(7월 16일부터 7월 26일)은 초장과 주경, 그리고 분지 수의 급격한 생장 이후 완만해지는 경향이 관찰되었다. 특히 Fig. 2에서와 같이 장마기부터 영양생장의 종료로 추정되는 시기까지의 일조시간(8시간 미만) 일수는 평야지가 22~29일, 중산간지가 26~35일 경과한 것으로 조사되었는데, 이는 22~34일의 단일조건 후 개화가 유도되었다는 선행 연구(Sohn and Kim, 2003)와 매우 유사하였다. 즉 Fig. 2와 3의 결과를 근거로 영양생장에서 생식생장으로 전환되는 시점에 적용된 8시간 미만의 일조시간이 단일조건(광주기성)에 영향을 주었을 것으로 추정되며, 추후 꽃향유의 개화를 유도하는 의존적 광주기와 파종시기에 따른 개화 특성에 관한 연구가 필요할 것으로 생각되었다.
재배지대와 유형별 개화 및 생육량 비교
꽃향유의 재배지대와 유형에 따른 개화 특성을 조사한 결과는 Table 2와 같다. 평야지와 중산간지 모두 비가림하우스에서 노지 대비 화서의 길이가 길고 화서경이 두꺼웠으며, 화서당 꽃수가 많은 편이었나 통계적 차이는 없었다. 화서폭은 평야지보다 중산간지가, 노지보다 비가림하우스에서 더 넓은 경향이었으며, 주당 화서수는 노지(평야지와 중산간지가 각각 807±88.1개, 620±48.0개) 대비 비가림하우스(각각 1,620±111.1개, 1,700±128.1개)에서 유의하게 높았다.
Table 2.
The effects of different altitudes and cultivation methods on flower characteristics at the late bloom stage of Elsholtzia splendens.
|
Different altitudes |
Cultivation methods |
Spike length (㎝) |
Spike width (㎜) |
Spike diameter (㎜) |
Number of spikes (per plant) |
Number of flowers (per spike) |
| Plains | Field | 6.70±0.146z | 1.20±0.028 | 1.45±0.033 | 807±88.1 | 213.8±11.15 |
| Plastic house | 7.42±0.208 | 1.22±0.029 | 1.65±0.045 | 1,620±111.1 | 241.7±9.74 | |
| Mid-mountain | Field | 6.90±0.145 | 1.31±0.037 | 1.57±0.041 | 620±48.0 | 210.9±14.79 |
| Plastic house | 7.96±0.170 | 1.47±0.033 | 1.89±0.069 | 1,763±98.7 | 269.6±13.75 | |
| Significance | ||||||
| Different altitudes (A) | NS | NS | NS | NS | NS | |
| Cultivation methods (B) | NS | NS | NS | * | NS | |
| A × B | NS | * | NS | NS | NS | |
수확기 생장과 수량 구성요소(Table 3) 중, 초장, 초폭, 주경, 분지수 및 생체중은 비가림하우스에서 노지 대비 생육량이 많았으나 대체로 재배지대에 따른 통계적 차이는 명확하지 않았다. 반면 건물률은 노지에서 비가림하우스에 비해 높은 경향이었다. 또한 중산간지에서 노지 대비 정유 추출량이 많은 편이었으나 통계적 차이는 없었으며, 폴리페놀 함량, ABTS free radical 소거능 등도 동일한 결과를 보였다(데이터 미제시).
Table 3.
The effects of different altitudes and cultivation methods on growth characteristics and yield at the late bloom stage of Elsholtzia splendens.
|
Different altitudes |
Cultivation method |
Plant length (㎝) |
Plant width (㎝) |
Stem diameter (㎜) |
Number of branchesz |
Fresh weight (g, per plant) |
Dry matter (%, per plant) |
Essential oil extraction (mL)y |
| Plains | Field | 94.7±2.62x | 85.3±3.73 | 14.3±0.25 | 163.5±3.40 | 1,066±68.2 | 31.7±19.9 | 1.95±0.226 |
| Plastic house | 143.2±5.52 | 108.7±2.44 | 22.4±0.96 | 222.9±8.90 | 2,191±189.8 | 25.3±50.8 | 1.86±0.201 | |
|
Mid- mountain | Field | 80.1±2.54 | 81.3±4.05 | 9.3±0.47 | 103.4±4.62 | 855±57.2 | 29.7±17.3 | 2.37±0.175 |
| Plastic house | 143.2±5.44 | 107.2±2.94 | 18.4±0.87 | 192.2±2.97 | 2,142±157.7 | 25.8±46.2 | 2.18±0.207 | |
| Significance | ||||||||
| Different altitudes (A) | NS | NS | * | NS | NS | NS | NS | |
| Cultivation methods (B) | * | * | * | NS | * | NS | NS | |
| A × B | NS | NS | NS | * | NS | * | NS | |
대표적인 단일 식물인 국화는 재배고도가 높을수록 비례적인 생장량과 수량을 나타내며(Afriani et al., 2022), 딸기의 경우 고랭지 생산 묘는 평난지에 비해 화아분화와 밀접한 관계가 있는 C/N율이 높고(Lee and Sugiyama, 1971), 자묘 생산에는 평난지의 온난한 기후가 유리하다(Lee et al., 2010). 꽃향유도 선행 연구들과 마찬가지로 생식생장을 나타내는 개화 특성은 중산간지 재배에서, 영양생장량을 나타내는 초장 등은 평야지 재배에서 높은 경향이었다.
결론적으로 자연적인 재배조건 하에서 꽃향유의 영양생장의 증가는 정식 후 59~69일경 완만해졌으며, 동 시기 광주기성의 영향으로 생식생장으로의 전환이 이루어져 9월 상순경 화뢰가 관찰된 것으로 판단되었다. 재배지대별 꽃향유의 화서 생장량 중 화서폭과 주당 화서수는 평야지보다 중산간지가 많은 경향이었고, 기본 식물체의 생장량은 주경, 분지수, 건물률을 제외하고는 통계적인 차이가 없었다. 화서를 포함한 재배유형별 생장량은 노지 재배 대비 비가림하우스 재배에서 비교적 많은 경향이었다. 따라서 중산간지와 평야지에서 경관 목적의 꽃향유를 재배할 시 화서폭과 주당 화서 수의 일부 차이는 있었으나 개화 및 생장 특성에 큰 차이를 보이지 않아 지대 구분없이 활용이 가능할 것으로 판단되었다. 또한 비가림하우스 대비 노지 재배에서는 본 실험의 초폭 조사 결과를 기준으로 약 21~24% 밀식재배가 필요할 것으로 생각되었다. 추가적으로 정유 추출량은 통계적 유의성은 없었으나 중산간지에서 평야지 대비 비교적 높은 결과를 나타내어 건조 및 추출방법, 유효성분 변화 등 관련 연구가 필요할 것으로 생각된다.
적 요
본 연구는 꽃향유(Elsholtzia splendens)의 재배지대(평야지, 중산간지)와 유형(노지, 비가림하우스 재배)에 따른 생육 및 개화 특성에 대해 구명하고자 수행하였다. 평야지와 중산간지는 각각 해발고도 12 m, 500 m에 위치한 시험포장을 선정하였으며, 재배지대와 유형별 재배관리는 동일한 방법을 적용하였다. 중산간지에서는 평야지 대비 화뢰 출현기(2~3일), 개화기(9일), 만개기(6~7일)가 빨랐으며, 재배유형 중 노지 재배에서 비가림하우스 재배보다 빠른 편이었다. 초장, 주경, 분지수는 정식 후 59~69일까지 급격히 증가한 후 완만해지는 경향을 보였다. 특히 장마기(6월 20일)부터 영양생장의 증가가 완만해지는 시기(정식 후 59~69일)까지의 일조시간(8시간 미만) 일수는 평야지와 중산간지가 각각 22~29일, 26~35일로 조사되었으며, 해당시기는 영양생장에서 생식생장으로 전환되는 시기인 것으로 추정되었다. 재배지대별 꽃향유의 화서 생장량은 평야지보다 중산간지에서 많은 경향이었고, 생육 특성은 주경, 분지수, 건물률을 제외하고는 통계적인 차이가 없었다. 또한 노지 재배 대비 비가림하우스 재배에서 개화량과 생장량이 많은 경향이었다. 결과적으로, 경관 목적의 꽃향유 재배 시 일부 개화량 차이는 인정되나 평야지와 중산간지 재배가 모두 가능할 것으로 판단된다. 이와 같은 결과는 꽃향유의 기상특성과 생육간의 관계를 이해하기 위한 기본적인 재배정보를 제공할 수 있을 것이다.
