

Introduction
Materials and Methods
Chemicals and reagents
Cell culture
Cell viability assay
NO production assay
DPPH assay
LC/MS analysis of chemicals in mulberry
Results
Effects of ME and MFE on cell viability in RAW 264.7 cells
Effects on NO production between mulberry and fermented mulberry in LPS-activated RAW 264.7 cells
Antioxidant activities of ME and MFE (DPPH)
UPLC analysis of chemicals in mulberry
Discussion
Introduction
Inflammation is a complex and essential physiological response to harmful external stimuli, including particulate matter, toxic compounds, bacteria, viruses, mechanical tissue injury, and aberrant autoimmune reactions (Wang et al., 2020a). A properly regulated acute inflammatory response contributes to tissue repair and healing. However, chronic inflammation can lead to persistent cellular and tissue damage, ultimately contributing to the development of various diseases, such as cancer, autoimmune disorders, cardiovascular diseases, diabetes, and neurodegenerative conditions (Wang et al., 2020b).
Inflammatory stimuli such as lipopolysaccharide (LPS) activate macrophages and trigger intracellular signaling pathways that result in the production of pro-inflammatory mediators, including tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1), interferons, prostaglandins, and nitric oxide (NO) (Kiemer et al., 2002). LPS, a major component of the outer membrane of Gram-negative bacteria, is one of the most common inducers of inflammatory responses. LPS-stimulated macrophages are particularly effective in producing a cascade of inflammatory cytokines and mediators (Wang et al., 2020b). In addition, LPS strongly induces the generation of NO, a key inflammatory mediator involved in immune regulation and tissue damage (Wang et al., 2020a).
In recent years, increasing attention has been directed toward the anti-inflammatory properties of phytochemicals derived from plant extracts used in traditional medicine, as inflammation is now recognized as a central factor in the pathogenesis of many diseases (Suriyaprom et al., 2023). Mulberry (Morus alba L.) is a flowering plant species belonging to the family Moraceae (Jung et al., 2019). During fruit development, mulberry fruits initially appear white, gradually change to pale yellow with pink margins, and eventually ripen to red. When fully mature, the fruits become deep purple and ultimately black (Maqsood et al., 2022).
The nutritional composition and antioxidant activity of mulberry fruits (Morus alba L.) vary depending on the stage of ripeness (Chen et al., 2022; Jelled et al., 2017). Mulberry fruits are rich in essential nutrients and exhibit diverse biological activities, including antimicrobial, anti-diabetic, and anti-hypertensive effects. They are also known for their ability to reduce blood glucose and cholesterol levels and to help prevent both acute and chronic diseases (Thabti et al., 2013).
Recent studies have demonstrated that fermentation of fruits with lactic acid bacteria (LAB) can significantly enhance the levels of bioactive compounds, including phenolics, flavonoids, and anthocyanins (Guan et al., 2024). In addition, probiotic LAB produces various volatile flavor compounds during fermentation, thereby improving the sensory and organoleptic properties of fermented products (Jiang et al., 2024). Therefore, the combination of LAB fermentation and mulberry fruit represents a promising strategy to enhance its potential health benefits.
Indeed, recent research has shown that fermentation of mulberry juice with Lactiplantibacillus strains, including L. plantarum, L. acidophilus, and L. paracasei, resulted in significant increases in total anthocyanin, phenolic, and flavonoid contents, along with enhanced antioxidant activity (Guan et al., 2024; Zhang et al., 2023). Accordingly, in the present study, mulberry fruit was fermented using Lactiplantibacillus plantarum, a strain widely employed in food fermentation. The objective of this study was to investigate the changes in antioxidant and anti-inflammatory activities of mulberry fruit extract before and after fermentation with L. plantarum.
Materials and Methods
Chemicals and reagents
The freeze-drying mulberry extracts (ME) of unripe mulberries (UM), semi ripe mulberries (SRM), and ripe mulberries (RM) were obtained from the National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea. Mulberry fermented extract (MFE) is a sample produced by fermenting UM, SRM, and RM with L. plantarum. L. plantarum was obtained from Korean Collection for Type Cultures (KCTC). Fetal bovine serum (FBS) and Dulbecco's modified Eagle Medium (DMEM) and penicillin/ streptomycin were purchased from Gibco. LPS and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma. Dimethyl sulfoxide (DMSO) was purchased from Samchun. 1,1‐diphenyl‐2‐picrylhy‐drazyl (DPPH) was supplied by Sigma. Rutin (MFDS-H-17-2-027) was purchased from Ministry of Food and Drug Safety of the Republic of Korea. Methanol was purchased from Thermo Fisher Scientific (Hampton, NH, USA). Acetonitrile and water were purchased from J.T. Baker (Phililpsburg, NJ, USA). Formic acid was purchased from Daejung (Siheung, Gyeonggido, Republic of Korea). All compounds and reagents were HPLC grade.
Cell culture
RAW 264.7 macrophage cells were cultured in DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin (100 U/mL penicillin and 100 µg/mL streptomycin) and maintained at 37℃ in a 5% CO2 incubator. The medium was replenished three times a week, and cells were subcultured without using trypsin.
Cell viability assay
Cell viability was determined by using MTT assay. Briefly, the cells were grown in 24-well plates at a density of 2×105 cells/well and incubated at 37℃ in a 5% CO2 incubator for 24 h. The media was changed and then treated with the medium supplemented with various concentrations of ME and MFE (5-500 µg/mL) and incubated at 37℃ for 24 h. Fifty microliters of MTT solution (5 ㎎/mL) was added and then cells were incubated for 4 h at 37℃. After removing the supernatant, the insoluble formazan product was dissolved in DMSO. Then, optical density of 96-well cultured plates was measured using Versamax microplate reader (Molecular Devices, CA, USA) at 540 ㎚.
NO production assay
NO formation was detected based on the accumulation of nitrite, an indicator of NO synthesis, in the culture medium via the Griess reaction. RAW 264.7 cells were plated at 2×105 cells/well in a 24-well and incubated at 37℃ in a 5% CO2 incubator for 24 h. The media were then changed and the cells were treated with 5 µL of non-toxic concentration of ME and MFE (5–500 µg/mL) for 4 h, then LPS at 1 µg/mL was added to stimulate inflammation. After 24 h of incubation, NO concentration was determined by measuring the amount of nitrite in the cell culture supernatant using the 100 µL of cell culture medium was mixed with 100 µL of Griess reagent (equal volumes of 1% (w/v) sulfanilamide in 5% (v/v) phosphoric acid and 0.1% (w/v) naphthyl ethylenediamine dihydrochloride), and the mixture was incubated at room temperature for 5 min. Absorbance was measured at 540 ㎚ wavelength using measurement at 540 ㎚ wavelength was obtained using the Versamax microplate reader (Molecular Devices, CA, USA).
DPPH assay
2,2‐diphenyl‐1 picrylhydrazyl (DPPH) radical scavenging activity was evaluated according to the method described by Sarikurkcu with slight modifications. Briefly, 20 µL of the sample solution with different concentrations was mixed with 100 µL of fresh methanolic DPPH solution (0.2 mM). After 30 min of incubation at room temperature, absorbance of the reaction mixture was measured using a Versa-max plate reader (Molecular devices) at a wavelength of 517 ㎚. L-ascorbic acid and Trolox were used as the reference compound. The scavenging rate (%) of DPPH free radicals was calculated according to a formula:
where A sample was the absorbance of 20 µL of sample solution and 180 µL of DPPH solution; A blank was the absorbance of 20 µL of 80% methanol solution and 180 µL of DPPH solution; and A control was the absorbance of 20 µL of ascorbic acid and 180 µL of DPPH solution. The DPPH radical scavenging activity (%) of the ME and MFE was determined as the percent drop in absorbance of the sample relative to the control.
LC/MS analysis of chemicals in mulberry
Ultra-Performance Liquid Chromatography was performed on the ACQUITY UPLC system (Waters, MA, USA) with an ACQUITY binary solvent manager pump (ACQ-BSM) and a ACQUITY PDA detector (ACQ-PDA). All extracts and compounds were analyzed under the following conditions: column, ACQUITY UPLC BEH C18 (2.1 × 100 ㎜, 1.7 μm; USA); mobile phase, 0.1% formic acid in distilled water (solvent system A), and 0.1% formic acid in acetonitrile (CH3CN, solvent system B) in a gradient mode (0.0-10.0 min 85-60% A; 10.0-10.1 min 60-0% A; 10.1-12.0 min 0-0% A; 12.0-12.1 min 0-85% A); sample injection volume, 3 μL; flow rate, 0.3 mL/min; column temperature, 40°C, UV wavelength, 320 ㎚.
Results
Effects of ME and MFE on cell viability in RAW 264.7 cells
The potential effects of ME and MFE on cell viability in RAW 264.7 cells were analyzed by the MTT assay after incubating cells for 24 h. To evaluate the effect of ME and MFE on the viability of RAW264.7 cells, MTT assay was performed. Results indicated that UM, SRM, RM (DW) and UM, SRM, RM (DW + L.p) at a concentration of up to 100 μg/mL had no significant toxic effects on the viability of RAW264.7 cells. The data showed that the viability of RAW264.7 cells treated with 5, 50, 100, 500 μg/mL ME and MFE for 24 h was not significantly different from that of the blank group (P > 0.05), indicating no cytotoxicity; therefore, subsequent experiments were conducted at concentrations of 5–100 μg/mL (Fig. 1). The results suggest that ME and MFE (5-100 μg/mL) has no cytotoxicity for RAW264.7 cells.
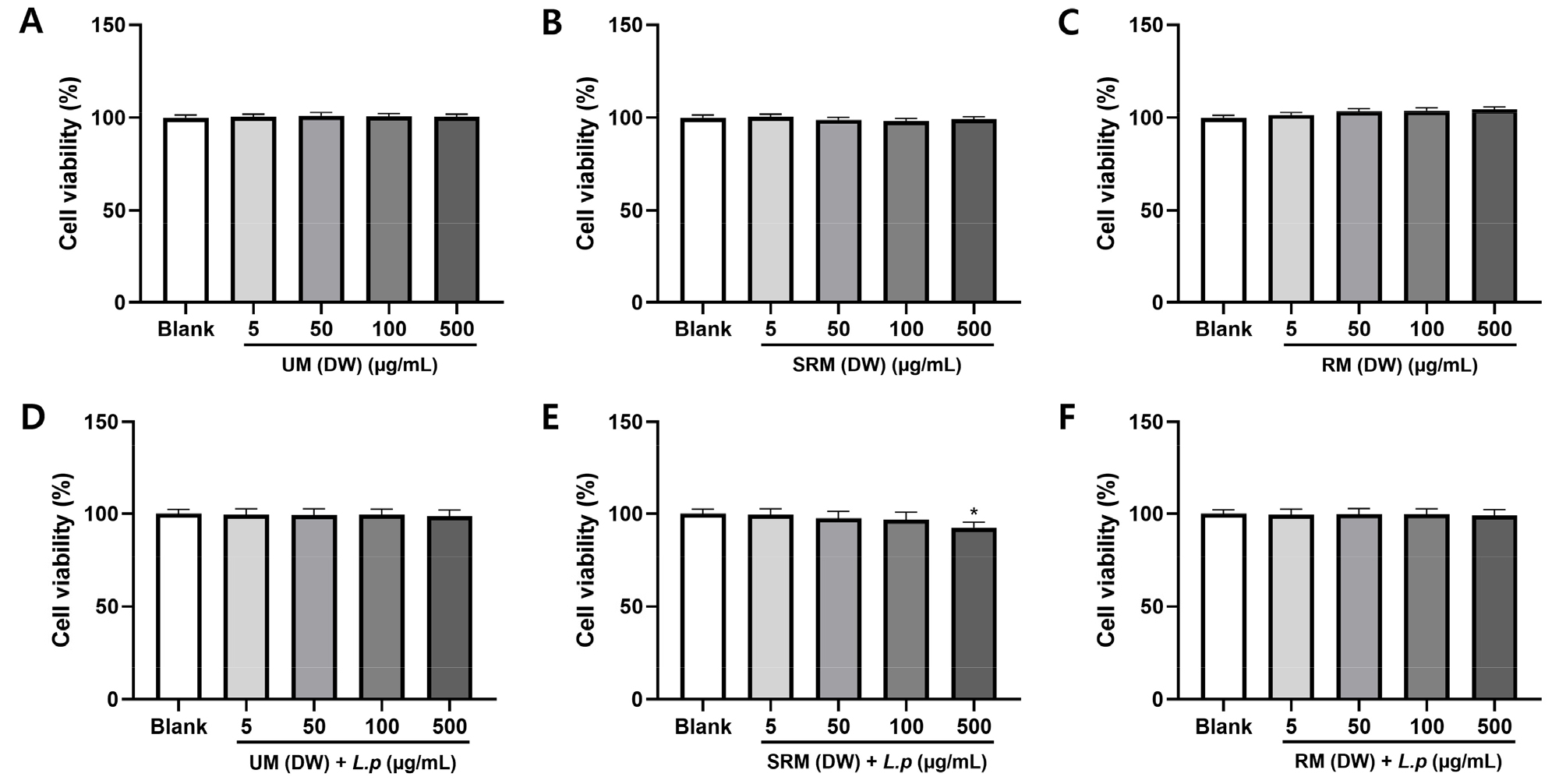
Fig. 1.
Effect of ME and MFE on cell viability in RAW264.7 cells. Cells were treated with different concentrations of UM, SRM, RM (5, 50, 100, 500 µg/mL) for 24 h. (A) RM (DW), (B) SRM (DW), (C) RM (DW), (D) UM (DW) + L.p (L. plantarum), (E) SRM (DW) + L.p, (F) RM (DW) + L.p. Data are presented as mean ± SD (one-way ANOVA), *p < 0.05 compared with sample-untreated cells.
Effects on NO production between mulberry and fermented mulberry in LPS-activated RAW 264.7 cells
In this study, NO production was measured in the culture medium to determine the anti-inflammatory activity of ME and MFE. We tested its effect on NO production in LPS-activated RAW 264.7 cells. As shown in Fig. 2, cells were treated with the ME and MFE before the LPS stimulation for the prevention of inflammation. All concentrations, except ME (SRM DW) 100 and 500 µg/mL, significantly reduced nitrite levels in LPS-activated RAW 264.7 cells. Considering the cytotoxic effects, the SRM (DW) + L.p group was evaluated at concentrations of 5–100 µg/mL. LPS exposure activated RAW264.7 cells inflammatory response, as NO secretion in the supernatants significantly enhanced after LPS stimulation for 24 h, and pre-treatment with various concentrations of ME and MFE in prior to LPS challenge notably attenuated the enhancement of these NO secretions. The NO production was higher in the LPS group than in the control group. ME and MFE was found more potent to inhibit LPS-induced NO generation (Fig. 2). These results indicated that ME and MFE exerts anti-inflammatory activity via the suppression of NO production in LPS-induced RAW264.7 cells. Therefore, it is suggested that ME and MFE exerts a protective effect on inflammation by inhibiting NO production in LPS-stimulated macrophages.
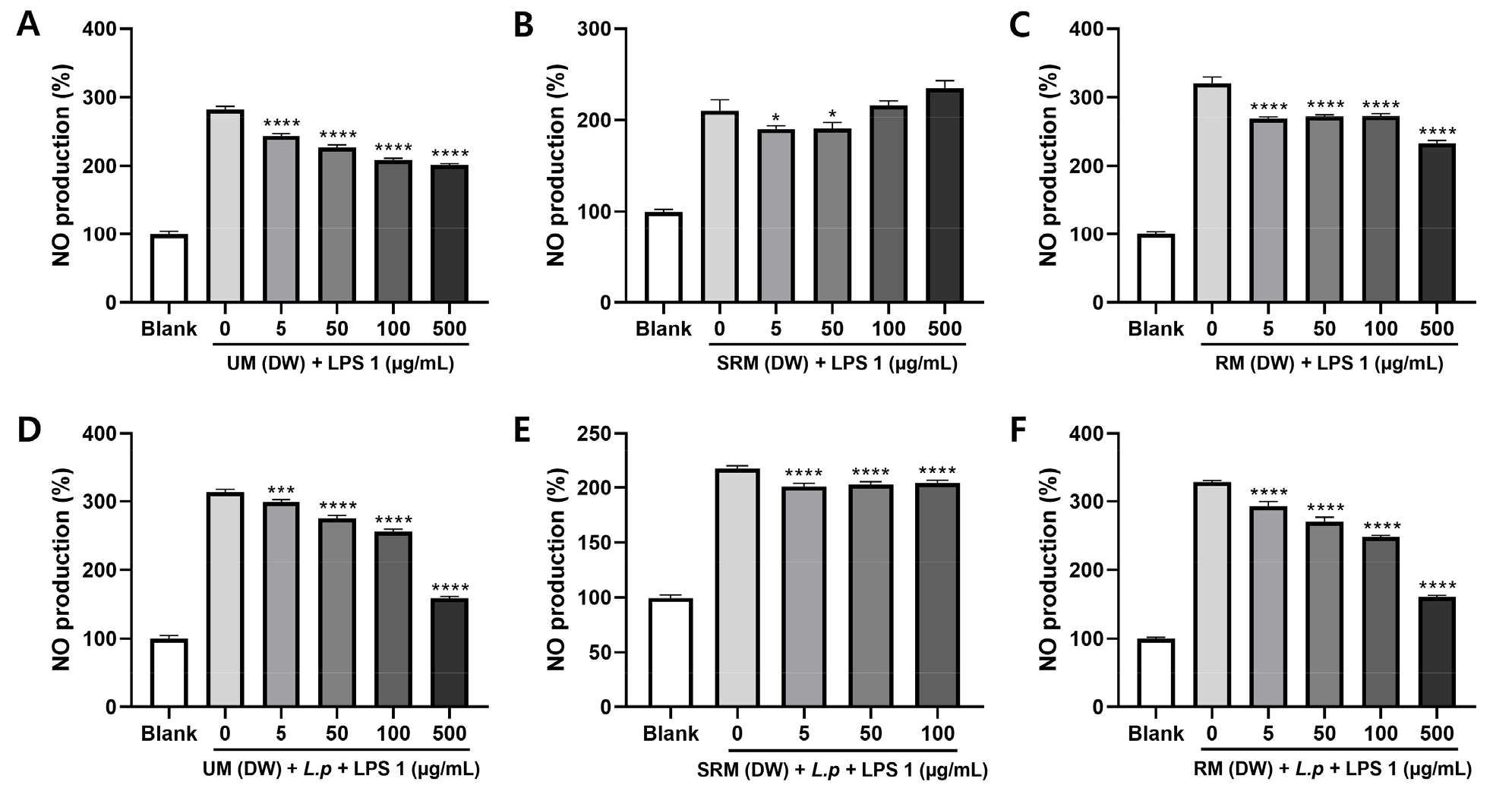
Fig. 2.
Effect of ME and MFE on NO production in RAW RAW264.7 cells. (A-F) Effects of UM, SRM and RM on NO production in RAW 264.7 mouse macrophages treated with LPS. (A) UM (DW), (B) SRM (DW), (C) RM (DW), (D) UM (DW) + L.p (L. plantarum), (E) SRM (DW) + L.p, (F) RM (DW) + L.p. Data are presented as mean ± SD (one-way ANOVA), * p < 0.05 compared to group treated with 1 µg/mL LPS alone.
Antioxidant activities of ME and MFE (DPPH)
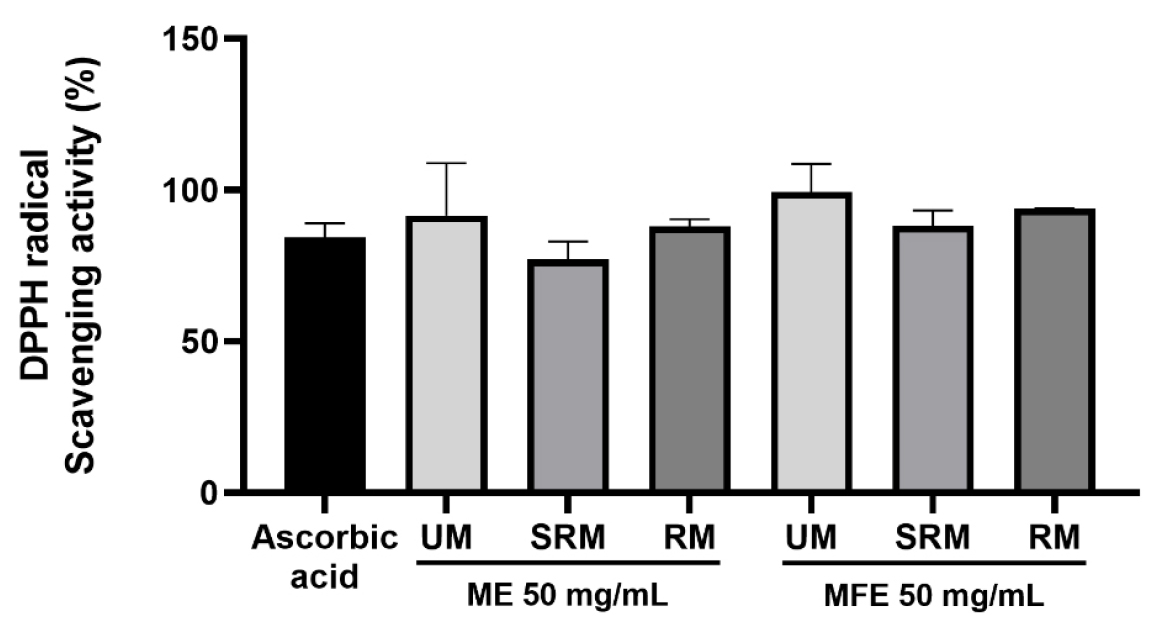
The antioxidant abilities of ME and MFE were firstly evaluated by measuring the DPPH radicals scavenging capacity. At first as shown in Fig. 3, ME and MFE exhibited obvious scavenging effect of DPPH radicals in a 50 ㎎/mL (50,000 µg/mL). The DPPH radical scavenging rates were as follows: the UM of ME exhibited a scavenging rate of 91.6%, the SRM showed 77%, and the RM demonstrated 88.1%. For MFE, the UM exhibited a scavenging rate of 99.3%, the SRM showed 88.2%, and the RM presented 93.9%. Collectively, MFE showed superior radical scavenging capacity compared to ME.
UPLC analysis of chemicals in mulberry
To examine the changes in components in ME and MFE, we conducted quantitative analysis by selecting rutin as an indicator chemical. The rutin peak was observed at 3.02 min (Fig. 4). The rutin contents were measured as follows: 2.64 ㎎/g for UM, 1.88 ㎎/g for SRM, and 1.11 ㎎/g for RM. Similarly, the rutin contents in the fermented samples were 2.49 ㎎/g for UM + L.p, 1.73 ㎎/g for SRM+ L.p, and 1.05 ㎎/g for RM + L.p. These results indicate that rutin content decreases as mulberry ripens, with no significant difference before and after fermentation. However, because rutin alone may not fully reflect the compositional changes induced by fermentation, further comprehensive analyses of other flavonoids, phenolic compounds, and fermentation-derived metabolites are warranted to better elucidate the chemical modifications occurring during fermentation.

Discussion
This study confirmed the potential of Morus alba fruit extract (ME) and fermented Morus alba fruit extract (MFE) as natural functional materials by evaluating their antioxidant and anti-inflammatory activities. In the DPPH radical scavenging assay, which was used to assess antioxidant activity, the MFE group at a concentration of 50 ㎎/mL exhibited an effect comparable to that of the positive control, ascorbic acid, demonstrating superior antioxidant activity compared with the ME group. To evaluate anti-inflammatory activity, the cytotoxicity of ME and MFE in RAW 264.7 macrophages was assessed using the MTT assay. The ME groups (UM, SRM, and RM) and the MFE groups (UM and RM) showed nearly 100% cell viability at concentrations below 500 μg/mL. Therefore, subsequent experiments were conducted at concentrations of 5, 50, 100, and 500 μg/mL. However, the MFE-SRM group exhibited cytotoxicity at 500 μg/mL; thus, further experiments for this group were performed at concentrations of 5, 50, and 100 μg/mL. Overall, these results demonstrate that L. plantarum-fermented mulberry fruit extracts exhibit enhanced anti-inflammatory and antioxidant activities compared with non-fermented mulberry fruit extracts, suggesting that fermentation is an effective approach to improving the biological functionality of mulberry fruits.
These findings are consistent with previous studies, such as the fermentation of hydroponic ginseng using Lactococcus lactis KC24, in which fermentation significantly enhanced antioxidant capacity and markedly reduced nitric oxide (NO) production (from 9.87 μM to 1.62 μM) (Chung et al., 2021). This observation supports our results, suggesting that fermentation of Morus alba (mulberry) may similarly exert NO-suppressive effects, potentially through increased bioavailability or biotransformation of antioxidant compounds.
Although rutin was selected as a representative marker compound in this study, no significant difference in rutin content was observed between ME and MFE. However, fermentation is known to induce extensive changes in the overall phytochemical composition rather than altering a single compound. Previous studies have reported that microbial fermentation can convert glycosylated flavonoids and bound polyphenols into their aglycone or low-molecular-weight phenolic forms through enzymatic reactions mediated by β-glucosidase and esterase, thereby enhancing biological activity and cellular uptake (Filannino et al., 2018; Hur et al., 2014; Yang et al., 2023). Therefore, the enhanced anti-inflammatory and antioxidant activities observed in MFE may be attributed to compositional changes or the generation of bioactive metabolites during the fermentation process rather than changes in rutin content alone.
Recent studies have demonstrated that inflammation plays a critical role in the progression of numerous chronic diseases, including diabetes, cancer, cardiovascular diseases, ocular disorders, arthritis, obesity, autoimmune diseases, and inflammatory bowel disease. The excessive production of free radicals from various biological and environmental sources often results from an imbalance between oxidants and endogenous antioxidant defenses, thereby contributing to the development of inflammation-related diseases (Arulselvan et al., 2016). Under normal physiological conditions, a balance is maintained between the generation of reactive oxygen species (ROS) or free radicals and the body’s endogenous antioxidant defense systems. However, disruption of this balance leads to oxidative stress, which can cause cellular and tissue damage and promote the progression of inflammatory and degenerative diseases (Birben et al., 2012).
The increased DPPH radical scavenging activity of MFE may be associated with its inhibitory effect on NO production. Antioxidant activity can attenuate intracellular oxidative stress, which plays a critical role in the induction of iNOS and excessive NO generation in LPS-stimulated macrophages (Han et al., 2001). Moreover, ROS readily interact with NO to form reactive nitrogen species such as peroxynitrite, contributing to redox imbalance and amplification of inflammatory responses (Pacher et al., 2007; Kim and Park, 2026). Therefore, the enhanced radical scavenging capacity of MFE may contribute to the maintenance of redox homeostasis, thereby limiting NO overproduction under inflammatory conditions.
Oxidative stress can damage essential cellular components, including DNA, proteins, and membrane lipids, potentially leading to cell dysfunction and death (Özcan et al., 2015). The inflammatory process is a major contributor to the induction of oxidative stress and the reduction of cellular antioxidant capacity (Khansari et al., 2009). Oxidative stress arises when the balance between pro-oxidants and antioxidants in the body is disrupted, often as a consequence of inflammation. This imbalance ultimately exerts deleterious effects on cells and tissues, resulting in oxidative damage.
Reactive oxygen species (ROS), which play a central role in oxidative injury, can directly interact with reactive nitrogen species (RNS), including nitric oxide (NO). These interactions influence the bioavailability and reactivity of NO and contribute significantly to the regulation and amplification of inflammatory responses (Wink et al., 2011). Nitric oxide (NO) plays a critical role in regulating cellular and physiological processes by inhibiting oxidative damage and protecting against cell death; however, excessive NO production can exert detrimental effects, including tissue injury and genetic mutations (Ohshima and Bartsch, 1994; Wink et al., 2001). Therefore, natural antioxidants that scavenge reactive oxygen species (ROS), including NO, and suppress oxidative stress are important candidates for the development of anti-inflammatory agents.
In this study, both Morus alba fruit extract (ME) and fermented Morus alba fruit extract (MFE) exhibited significant anti-inflammatory activity, and their antioxidant properties were also clearly demonstrated. Macrophages are key components of the innate immune system and play pivotal roles in inflammatory responses through antigen presentation, phagocytosis, and the production of various cytokines and growth factors (Jang et al., 2022; Lee and Choi, 2018).
Fermentation is a biological process that improves the chemical composition of foods, modulates the gut microbiota, and generates beneficial compounds through microbial activity. Historically, fermentation has been widely used in food processing, and numerous studies have demonstrated the health-promoting effects of fermented foods, including enhanced nutrient bioavailability and improved physiological functions of food components. In recent years, research has increasingly focused on applying fermentation techniques to natural products to achieve enhanced and more diverse biological activities (Park and Mannaa, 2025). For example, fermentation using Lactiplantibacillus species has been reported to improve the nutritional quality of fruit juices. In one study, Cerasus humilis juice was fermented with two commercial probiotic strains, Lactobacillus acidophilus and Lactobacillus plantarum, resulting in enhanced chemical properties, including increased total antioxidant capacity (TAOC) and higher viable cell counts. Moreover, fermentation enhanced the in vitro antioxidant activity of the juice following simulated gastrointestinal digestion and induced significant alterations in the gut microbiota composition of the fermented product. Furthermore, co-fermentation with Lactobacillus acidophilus and Lactobacillus plantarum significantly enhanced the antioxidant properties of the juice after digestion and increased the abundance of probiotic populations capable of modulating the gut microbiota (Li et al., 2023).
In addition to improving the nutritional value of the juice, this study demonstrated that fermentation induces notable alterations in gut microbiota composition (Li et al., 2023). Similarly, another study reported that fermented Lentinus edodes liquid exhibited superior nutritional, functional, and sensory properties compared with its unfermented counterpart (Zhang et al., 2022).
Moreover, fermented products involving Lactiplantibacillus plantarum have been shown to exert beneficial effects in the management of inflammatory diseases, including colitis and hypertension (Levit et al., 2017; Yi et al., 2024).
Therefore, we evaluated the antioxidant effects of Morus alba fruit extract (ME) and fermented Morus alba fruit extract (MFE). Our results revealed a trend toward increased radical scavenging activity in MFE compared with ME, suggesting that compositional changes induced by fermentation or metabolites generated during the fermentation process may act as active constituents that enhance DPPH radical scavenging activity. Furthermore, we observed differential biological effects between untreated ME and MFE fermented with Lactiplantibacillus plantarum (L.P.) in LPS-stimulated RAW 264.7 macrophages.
A limitation of this study is that we did not evaluate the effects of ME and MFE on the production of pro-inflammatory cytokines, such as interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6), following LPS stimulation. However, previous studies have reported that fermented mulberry leaf extract significantly reduced the expression of IL-1β, demonstrating notable therapeutic and preventive effects against intestinal mucositis (Oh et al., 2017).
These findings suggest that Morus alba fruit extract, especially MFE, may have potential anti-inflammatory effects. In conclusion, our results indicate that MFE exerts anti-inflammatory activity, at least in part, through the suppression of NO production. Nevertheless, further mechanistic studies are required to elucidate the effects of MFE on inflammatory signaling pathways and the expression of key inflammatory mediators, including inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), TNF-α, IL-1β, and IL-6.
