

서 언
유채(Brassica napus L.; AACC, 2n=38)는 십자화과(Brassicacea) 배추속(Brassica)에 속하는 이질4배체(allotetraploid)로 순무(Brassica rapa L.; AA, 2n=20)와 양배추(Brassica oleracea L.; CC, 2n=18)의 자연 종간 교잡에 의해 유래된 것으로 알려져 있다(Iniguez-Luy and Federico, 2011). 현재 재배되고 있는 유채는 저온 조건이 없는 온화한 기후에서 파종 연도에 꽃을 피우는 봄형(spring type), 중국 북부와 같이 겨울철 기온이 급격하지 않은 지역에서 재배되는 반겨울형(semi-winter type), 그리고 북유럽과 같이 개화를 위해 저온 기간이 의무적으로 필요한 겨울형(winter type) 등 세 가지 유형으로 구분된다(Lee et al., 2014a; Leijten et al., 2018).
우리나라에서 유채는 재배면적이 1998년 정부 수매 중단 이전까지 꾸준히 증가하였으나 이후 수매 중단과 외국산 올리브유, 대두유 및 옥수수유 등의 대체 식용유 수입의 급격한 증가로 유채 재배면적이 지속적으로 감소되었다(Jung et al., 2007). 국내 유채 재배는 주로 10월 중순 파종하여 이듬해 4월 중순-하순에 개화성기가 되고, 6월 중순에 수확하는 가을 재배를 시행하는 동계작물로서 타 식량작물과 경합을 피할 수 있으며, 전초를 청예사료로 이용이 가능한 장점이 있다(Kim et al., 2015b; Trethewey, 2012).
현재는 주로 봄철 지역 축제의 대표 경관작물로 재배되며, 최근에는 소비자의 안전하고 건강한 식품에 대한 관심 증가로 일반 식용유를 대체할 수 있는 고급 유채유에 대한 수요가 증가되고 있어 정부의 경관작물직불제에 따른 유채 재배와 더불어 유채유 생산을 통해 농업 부가가치를 높일 수 있는 소득 작물로서 그 가치가 부각되고 있다(Kim et al., 2007).
유채 종자의 식물성 기름은 팜유 및 대두유와 함께 세계적으로 중요한 유지 작물 중 하나이다(Chew, 2020; FAO, 2014). 특히 유채는 저에루스산 유채(LEAR, Low Erucic Acid Rapeseed)와 고에루스산 유채(HEAR, High Erucic Acid Rapeseed)로 분류할 수 있는데, 다른 십자화과 식물보다 많은 에루스산(C22:1)을 함유하고 있는 고에루스산 유채는 최대 66%까지 함유할 수 있다(Rui et al., 2014). 추출한 에루스산은 디젤 연료, 화학 원료, 방수제 및 윤활제 등 산업 응용 분야에 광범위하게 이용되어 많은 연구자들이 에루스산의 산업적 활용에 대한 연구를 진행하고 있다(Mcvetty and Duncan, 2015; Qi and Wnag, 2009; Taylor et al., 2011; Wu et al., 2007).
식품 산업에서 에루스산은 장기간 섭취 시 심근섬유증, 심근병증, 심장 근육 및 신장의 지방 침착 등 여러 부작용이 보고되었다(Bremer and Norum, 1982; Flatmark et al., 1983; Kramer et al., 1992; Reyes et al., 2010; Vles et al., 1978). 이에 따라, 유채 종자의 에루스산 함량을 낮춘 품종 육성으로 저에루스산 유채가 개발되었고(Lühs and Friedt, 1994; Przybylski and Mag, 2002), 현재 중국은 국가 표준 GB/T1536-2004에 따라 에루스산 함량이 3% 미만인 유채를 저에루스산 유채로 간주하고 유럽연합은 규정 2019/1870에서 소비자에게 제공되는 식물성 기름 및 지방에 함유된 에루스산의 최대 함량을 2% 이하로 규정하였다(Wang et al., 2022). 우리나라도 2023년 식품의약품안전처 고시 제2023-56호에 유채유의 에루스산 기준을 신설하여 함량 2% 이하로 규정하고 있다.
식용 유채유의 품질은 주로 올레인산(oleic acid, C18:1), 리놀레산(linoleic acid, C18:2), 리놀렌산(linolenic acid, C18:3)의 세 가지 주요 불포화지방산의 비율에 의해 결정된다(Gillingham et al., 2011; Micha and Mozaffarian, 2009). 과거에는 리놀레산과 리놀렌산이 인체의 필수 지방산으로 간주되었으나(Kim et al., 2013) 최근 연구로 올레인산이 더 중요한 필수 지방산인 것으로 보고되었다(Picklo and Murphy, 2016). 현재 전 세계적으로 재배되는 대부분의 유채는 에루스산 함량이 낮고 약 55∼65% 수준의 올레인산이 함유된 품종이다(Merrill et al., 2008; Zhao et al., 2019). 올레인산은 산화안정성이 높고 저온유동성이 우수한 단일불포화지방산으로 높은 활용도와 장기 저장이 가능하며, 섭취 시 혈당 조절, 심혈관 보호, 체지방 감소 등 건강상의 이점으로 대다수의 소비자들이 선호한다(Chang et al., 2022; Galassetti and Pontello, 2006; Liu et al., 2016; Petukhov et al., 1999; Salar-García et al., 2016).
국내에서도 종자에 에루스산을 함유하지 않은 무에루스산의 조숙형 ‘중모7001’과 만숙형 ‘중모7002’와 같은 우수 품종을 개발하였고, 최근에는 고올레인산을 함유하면서 도복과 균핵병에 강한 ‘유려’ 품종을 개발하는 등 꾸준한 품종 개발 연구로 유채 재배 활성화 및 유채유 생산을 촉진하고 있다(Jeong et al., 2024; Kim et al., 2014; Kim et al., 2015a). 국내 유채 재배 시 기계화가 가능한 논에서의 재배가 노동력 및 생산비용 절감 등으로 유리한데, 유채 파종기와 벼 수확기, 유채 수확기와 벼 이앙기가 중첩되어 이모작에 다소 어려움이 따른다. 이에 따라, 국내에서 파종시기 설정 연구, 파종시기에 따른 지방산 조성 변화 연구, 봄 재배 적합 품종 선발 연구, 기계이식과 직파 간 수량성 비교 연구 등 여러 연구들이 보고되어 있다(Kim et al., 2015b; Kwon et al., 2021; Lee et al., 2014a; Lee et al., 2014b; Lee et al., 2021). 연구 결과는 유채의 종자 수량성과 지방산 조성 구성 측면에서 파종 날짜가 늦어지면 수량이 감소하고 지방산 구성 비율이 변화한다고 보고하였는데, 이는 일반적으로 늦은 파종이 생육 기간 단축과 낮은 온도 및 기타 요인에 따른 스트레스로 인해 수확 지수가 낮아지고 기름 함량이 감소하며, 그 구성 또한 변화된다는 국외의 연구 결과(Kirkegaard et al., 2016; Ozer et al., 2003; Weymann et al., 2015; Xie et al., 2023)와 동일하였다.
본 연구는 우리나라 농업 작부체계 상황과 종자 생산성 및 지방산 조성을 고려하여 유채 경관 활용, 종자 생산, 유채유 활용 등 목적에 최적화된 유채 품종을 육성하기 위한 육종 소재로서 유전자원의 특성을 평가하고, 형질 간 통계분석으로 유용 자원을 분류하고자 한다(Kim et al, 2024; Lee et al, 2023; Yu et al, 2022). 유전자원의 형질 다양성 연구는 자원의 특성을 파악하여 유용 형질 발굴, 보존, 활용 및 관리를 위한 기초자료를 수집하는 중요한 분야로 계통분류, 전통육종, 분자육종 등 농업 여건에 적합한 신품종 개발에 적극 활용할 수 있는 연구이다(Cho et al., 2009; Jarvis et al., 2010; Rommens, 2007; Tolbert et al., 1979). 따라서, 본 연구의 목적은 유채 유전자원 형질의 다양성을 조사하고, 각 형질 간 상호관계를 분석하여 유사한 자원 간 군집화(clustering)를 통해 집단을 대표하는 특이적 형질을 도출하여 최적 품종 개발을 위한 교배 집단 선발의 기초자료를 수집하고자 한다.
재료 및 방법
실험재료
본 연구의 시험재료 유채(Brassica napus) 유전자원은 농촌진흥청 농업유전자원센터에서 보유하고 있는 유전자원 129종을 분양받아 이용하였다. 우리나라를 포함한 22개국에서 수집된 시험 유전자원은 국가등록 유전자원 목록에 부여된 명칭인 IT번호로 구분하였고, 원산지를 표기하였다(Table 1).
Table 1.
Information of the 129 Brassica napus genetic resources.
| # | IT No. | Name | Originz | # | IT No. | Name | Originz |
| 1 | 135171 | PI 311728 | POL | 66 | 279155 | BNW. 1 61/83 | DEU |
| 2 | 265048 | Hγ600-11-1-148-1-1 | KOR | 67 | 279156 | BNW. 1 62/83 | DEU |
| 3 | 279088 | Ames 15652 | USA | 68 | 279157 | Elena | DEU |
| 4 | 279089 | Ames 15653 | USA | 69 | 279158 | IR-2 | HUN |
| 5 | 279090 | Ames 15654 | USA | 70 | 279159 | Librador | DEU |
| 6 | 279091 | Ames 18935 | GBR | 71 | 279160 | Lindore | DEU |
| 7 | 279092 | Siberian | USA | 72 | 279161 | Linglandor | DEU |
| 8 | 279093 | Siberian | USA | 73 | 279162 | Liratrop | DEU |
| 9 | 279094 | Ames 6099 | UNK | 74 | 279163 | Liropa | DEU |
| 10 | 279095 | Ames 6102 | FRA | 75 | 279164 | Santana | DEU |
| 11 | 279096 | PI 184453 | DEU | 76 | 279165 | Ujfertodi | HUN |
| 12 | 279098 | PI 458607 | NZL | 77 | 279166 | Winfield | CAN |
| 13 | 279101 | PI 458918 | FRA | 78 | 279167 | Rebel | USA |
| 14 | 279102 | PI 458921 | FRA | 79 | 279168 | Cathy | USA |
| 15 | 279103 | PI 458922 | FRA | 80 | 279169 | TRI-Bridger | USA |
| 16 | 279104 | PI 458923 | FRA | 81 | 279170 | Delta | SWE |
| 17 | 279105 | PI 458924 | FRA | 82 | 279171 | Corvette | GBR |
| 18 | 279106 | PI 458925 | FRA | 83 | 279172 | KS 3579 | USA |
| 19 | 279107 | PI 458930 | CAN | 84 | 279173 | PI 597828 | DZA |
| 20 | 279108 | PI 458935 | SWE | 85 | 279174 | Global | SWE |
| 21 | 279109 | PI 458936 | SWE | 86 | 279175 | Topas | SWE |
| 22 | 279110 | PI 458937 | SWE | 87 | 279176 | Glacier | SWE |
| 23 | 279111 | PI 458939 | JPN | 88 | 279177 | Crystal | SWE |
| 24 | 279112 | PI 458940 | JPN | 89 | 279178 | Liradonna | USA |
| 25 | 279113 | PI 458944 | DEU | 90 | 279179 | PI 601504 | SWE |
| 26 | 279114 | PI 458945 | DEU | 91 | 279180 | AR91004 | USA |
| 27 | 279115 | PI 458946 | DEU | 92 | 279181 | Wichita | USA |
| 28 | 279116 | PI 458947 | DEU | 93 | 279182 | Abilene | USA |
| 29 | 279117 | PI 458948 | DEU | 94 | 279183 | PI 633118 | USA |
| 30 | 279118 | PI 458949 | DEU | 95 | 279185 | MAR | POL |
| 32 | 279120 | PI 458951 | DEU | 97 | 279188 | PI 633124 | SWE |
| 33 | 279121 | PI 458952 | DEU | 98 | 279189 | Sumner | USA |
| 34 | 279122 | PI 458953 | DEU | 99 | 279190 | PI 649126 | CAN |
| 35 | 279123 | PI 458954 | DEU | 100 | 279191 | PI 649127 | CAN |
| 36 | 279124 | PI 458955 | DEU | 101 | 279193 | PI 649129 | CAN |
| 37 | 279125 | PI 458956 | DEU | 102 | 279194 | PI 649130 | SWE |
| 38 | 279126 | PI 458957 | DEU | 103 | 279195 | Bronowski | POL |
| 39 | 279127 | PI 458958 | DEU | 104 | 279196 | Mazowiecki | POL |
| 40 | 279128 | PI 458959 | DEU | 105 | 279199 | Helga | DEU |
| 41 | 279129 | PI 458964 | NZL | 106 | 279200 | Baraska | DEU |
| 42 | 279130 | PI 458965 | NZL | 107 | 279202 | PI 649148 | ALB |
| 43 | 279131 | PI 458967 | FRA | 108 | 280312 | TN 190 | CHN |
| 44 | 279132 | PI 458968 | FRA | 109 | 280313 | TN 193 | CHN |
| 45 | 279133 | PI 458969 | FRA | 110 | 280314 | TN 194 | CHN |
| 46 | 279134 | PI 458970 | FRA | 111 | 280315 | TN 196 | CHN |
| 47 | 279135 | PI 458979 | USA | 112 | 280316 | TN 197 | CHN |
| 48 | 279136 | PI 458980 | USA | 113 | 280317 | TN 198 | CHN |
| 49 | 279137 | PI 469791 | FRA | 114 | 297368 | Red Russian | USA |
| 50 | 279138 | PI 469793 | FRA | 115 | 297524 | TN 050 | CHN |
| 51 | 279139 | PI 469794 | FRA | 116 | 298993 | Nγ600-21-1-641-1-1 | KOR |
| 52 | 279140 | PI 469795 | FRA | 117 | 299358 | TN 191 | CHN |
| 53 | 279141 | PI 469796 | FRA | 118 | 299359 | TN 195 | CHN |
| 54 | 279142 | PI 469797 | FRA | 119 | 299360 | TN 200 | CHN |
| 55 | 279143 | PI 469798 | FRA | 120 | 299361 | TN 201 | CHN |
| 56 | 279144 | PI 469799 | FRA | 121 | 299362 | TN 202 | CHN |
| 57 | 279145 | PI 469800 | FRA | 122 | 314692 | YP 26046 | CHN |
| 58 | 279146 | PI 469803 | DEU | 123 | 314693 | YP 26049 | CHN |
| 59 | 279147 | PI 469804 | DEU | 124 | 314695 | YP 26051 | CHN |
| 60 | 279148 | PI 470004 | POL | 125 | 314696 | YP 26052 | CHN |
| 61 | 279149 | PI 470005 | POL | 126 | 314697 | YP 26053 | CHN |
| 62 | 279150 | PI 470006 | POL | 127 | 314698 | YP 27106 | CHN |
| 63 | 279151 | PI 470007 | POL | 128 | 314699 | YP 27107 | CHN |
| 64 | 279152 | PI 470020 | ROU | 129 | 314700 | YP 27108 | CHN |
| 65 | 279154 | AR-256 | RUS |
zPOL, Poland; KOR, Korea; USA, United States; GBR, United Kingdom; UNK, Unknown; FRA, France; DEU, Germany; NZL, New Zealand; CAN, Canada; SWE, Sweden; JPN, Japan; ROU, Romania; HUN, Hungary; DZA, Algeria; ITA, Italy; ALB, Albania; CHN, China; MNG, Mongolia; TUR, Turkey; PHL, Philippines; SRB, Serbia; RUS, Russia.
재배 방법 및 농업 특성 조사
농촌진흥청 국립식량과학원 바이오에너지작물연구소 전작 포장(무안, 34°58´N 126°27´E, 해발 16 m)에서 재식거리 60 ㎝ × 30 ㎝로 10월 하순 파종하였다. 시비는 질소의 경우 기비 5 ㎏/10a와 2월 하순 생육재생기에 추비로 10 ㎏/10a 실시하였고, 인산과 가리는 P2O5 : K2O = 8 : 8 ㎏/10a로 전량 기비 실시하였다. 재배는 농촌진흥청 표준재배법에 의거하여 수행하였고, 농업 특성 조사는 농업 과학 기술 조사 분석 기준(RDA, 2012)에 준하여 개화시(DF, Days to Flowering), 경장(PH, Plant Height), 수장(MBL, Main Branch Length), 분지수(NB, Number of Branches), 수당협수(NP, Number of Pods per plant), 협장(PL, Pod Length), 종자수(NS, Number of Seeds per pod), 배주수(NO, Number of Ovules), 천립중(TSW, 1000-Seed Weight) 등 주요 농업적 형질을 조사하였다. 결실률(SSR, Seed Set Rate)은 배주수와 종자수로 산출하였다.
지방산 분석
종자 시료 0.5 g을 막자사발에서 마쇄한 후 pear-shaped flask에 넣고 99.5% methyl alcohol (CH3OH; DaeJung Chemical & Metals Co. Ltd, Korea) 15 mL과 sodium methoxide 30 wt% solution in methanol (Samchun Chemicals, Korea) 1 mL을 첨가하여 100℃에서 2시간 동안 methylation시켰다. 반응물을 상온에서 냉각시킨 후 15 mL n-hexane (Burdick & Jackson, USA)을 첨가하여 혼합하고 상층액을 필터링하여 Gas Chromatography (Agilent 7890A, Agilent technologies, USA)를 이용하여 지방산 조성을 분석하였다. 분석조건에 컬럼은 HP-Innowax (30 m × 0.32 ㎜ × 0.25 ㎛)를 사용하였으며, 검출기는 Flame ionization detector (FID)로 250℃에서 사용하였다. 주입부(Inlet)는 200℃로 사용하였으며, spilit mode injector는 10:1의 spilit mode로 하였다. 오븐 온도는 100℃에서 1분간 유지한 후 분당 8℃씩 250℃까지 상승시키고, 250℃에서 10분간 유지하여 각 peak의 면적을 상대적인 백분율로 나타내었다. 지방산 조성은 팔미트산(Palmitic acid), 스테아르산(Stearic acid), 올레인산(Oleic acid), 리놀레산(Linoleic acid), 리놀렌산(Linolenic acid), 에이코젠산(Eicosenic acid), 에루스산(Erucic acid) 등의 함량을 조사하였다.
통계분석
본 연구의 통계분석은 R studio software (www.-r.project.org, v.4.4.1., Vienna, Austria)를 사용하였다. 상관관계 분석은 피어슨 상관 계수(Pearson Correlation Coefficient, PCC)로 유의성을 확인하였고, 유채 유전자원의 형질 요인 간 표준화 주성분 분석(Principal Component Analysis, PCA)을 수행하여 유전자원의 형질별 분포와 주성분의 기여도를 분석하였다. 군집분석은 위계적 군집화 기법으로 유클리드 거리(Euclidean distance) 계산법에 기반하여 최장 연결법(Complete linkage method)으로 군집화하였다.
결과 및 고찰
유채 유전자원의 생육 특성 변이
유채 유전자원 129점에 대한 주요 농업적 형질 및 지방산 조성 특성의 평균, 표준편차 및 변이 계수와 각 특성의 최소값 및 최대값을 나타내는 유전자원을 확인하였다(Table 2).
개화소요일수는 최소 137일(IT279089, 미국) ∼ 최대 188일(IT279092, 미국)의 범위로 평균 172.4일이었다. 초장은 79.3 ㎝ (IT279146, 독일) ∼ 198.7 ㎝ (IT279162, 독일)의 범위로 평균 146.6 ㎝였고, 수장은 30.7 ㎝ (IT314698, 중국) ∼ 83.0 ㎝ (IT279098, 뉴질랜드)의 범위로 평균 49.9 ㎝였다. 분지수는 2.3 개(IT280317, 중국) ∼ 17.7 개(IT279107, 캐나다)의 범위로 평균 9.5 개였고, 수당협수는 18.3 개(IT314699, 중국) ∼ 103.0 개(IT279161, 독일)의 범위로 평균 60.7 개였다. 협장은 3.8 ㎝ (IT314700, 중국) ∼ 8.2 ㎝ (IT279164, 독일)의 범위로 분포하였고, 평균은 6.1 ㎝였다. 종자수는 13.3 개(IT279151, 폴란드) ∼ 32.2 개(IT279171, 영국)의 범위로 평균 24.2 개였고, 배주수는 15.6 개(IT314700, 중국) ∼ 36.5 개(IT279171, 영국)의 범위로 평균 27.7 개였다. 결실률은 73.1% (IT279165, 헝가리) ∼ 97.1% (IT314693, 중국)의 범위로 평균 87.2% 수준이었고, 천립중은 2.3 g (IT279186, 이탈리아) ∼ 6.4 g (IT279151, 폴란드)의 범위로 평균 4.1 g이었다.
지방산 조성으로 팔미트산(C16:0)은 최소 2.4% (IT279202, 알바니아) ∼ 최대 6.1% (IT279131, 프랑스)의 범위로 평균 4.2% 수준이었고, 스테아르산(C18:0)은 0.5% (IT279202, 알바니아) ∼ 1.6% (IT265048, 한국)의 범위로 평균 0.9% 수준이었다. 올레인산(C18:1)은 9.7% (IT279186, 이탈리아) ∼ 70.4% (IT279125, 독일)의 범위로 평균 43.9%였고, 리놀레산(C18:2)은 11.0% (IT280313, 중국) ∼ 27.1% (IT279126, 독일)의 범위로 평균 18.5%였다. 리놀렌산(C18:3)은 4.8% (IT279200, 독일) ∼ 15.6% (IT314698, 중국)의 범위로 평균 8.7%였고, 에이코젠산(C20:1)은 0.9% (IT279117, 독일) ∼ 18.4% (IT280317, 중국)의 범위로 평균 5.3% 수준이었고, 에루스산(C22:1)은 0% (IT279120, 독일) ∼ 51.9% (IT299362, 중국)의 범위로 평균 17.0% 수준이었으며, 그 외 지방산은 0 ∼ 6.6% 수준으로 평균 1.6% 가량 포함하였다.
Table 2.
Basic statistical data and information for agronomic traits and seed fatty acids content in 129 Brassica napus genetic resources.
| Statistics | Mean | SDp |
CVo (%) | Minimum | Maximum | ||||
| Value | IT No. | Origin | Value | IT No. | Origin | ||||
|
DFz (Days) | 172.4 | 8.43 | 0.05 | 137.0 | 279089 | United States | 188.0 | 279092 | United States |
|
PHy (㎝) | 146.6 | 21.72 | 0.15 | 79.3 | 279146 | Germany | 198.7 | 279162 | Germany |
|
MBLx (㎝) | 49.9 | 8.90 | 0.18 | 30.7 | 314698 | China | 83.0 | 279098 | New Zealand |
|
NBw (no.) | 9.5 | 3.04 | 0.32 | 2.3 | 280317 | China | 17.7 | 279107 | Canada |
|
NPv (no.) | 60.7 | 13.85 | 0.23 | 18.3 | 314699 | China | 103.0 | 279161 | Germany |
|
PLu (㎝) | 6.1 | 0.88 | 0.14 | 3.8 | 314700 | China | 8.2 | 279164 | Germany |
|
NSt (no.) | 24.2 | 3.60 | 0.15 | 13.3 | 279151 | Poland | 32.2 | 279171 | United Kingdom |
|
NOs (no.) | 27.7 | 3.62 | 0.13 | 15.6 | 314700 | China | 36.5 | 279171 | United Kingdom |
|
SSRr (%) | 87.2 | 5.05 | 0.06 | 73.1 | 279165 | Hungary | 97.1 | 314693 | China |
|
TSWq (g) | 4.1 | 0.74 | 0.18 | 2.3 | 279186 | Italy | 6.4 | 279151 | Poland |
|
Palmitic (C16:0, %) | 4.2 | 0.71 | 0.17 | 2.4 | 279202 | Albania | 6.1 | 279131 | France |
|
Stearic (C18:0, %) | 0.9 | 0.23 | 0.24 | 0.5 | 279202 | Albania | 1.6 | 265048 | Korea |
|
Oleic (C18:1, %) | 43.9 | 21.91 | 0.50 | 9.7 | 279186 | Italy | 70.4 | 279125 | Germany |
|
Linoleic (C18:2, %) | 18.5 | 3.44 | 0.19 | 11.0 | 280313 | China | 27.1 | 279126 | Germany |
|
Linolenic (C18:3, %) | 8.7 | 1.51 | 0.17 | 4.8 | 279200 | Germany | 15.6 | 314698 | China |
|
Eicosenic (C20:1, %) | 5.3 | 4.68 | 0.89 | 0.9 | 279117 | Germany | 18.4 | 280317 | China |
|
Erucic (C22:1, %) | 17.0 | 19.81 | 1.16 | 0.0 | 279120 | Germany | 51.9 | 299362 | China |
|
Others (%) | 1.6 | 1.41 | 0.88 | 0.0 | 279174 | Sweden | 6.6 | 314698 | China |
상관분석
유채 유전자원을 육종 소재로 이용하기 위한 선발에 필요한 적정 기준 설정을 위해 10가지 주요 농업적 형질과 8종의 지방산 조성 분석 결과로 상관관계를 조사하였다(Table 3). 작물의 재배기간과 밀접한 형질인 개화소요일수는 초장(r=0.633***)과 높은 정의 상관관계를 보였고, 수장(r=0.175*) 및 천립중(r=0.198*)과도 유의한 상관성을 보였다. 유채의 파종에서 개화까지 소요되는 개화소요일수가 많을수록 길어진 영양생장 기간에 따라 초장, 수장 및 천립중도 증가되기 때문인 것으로, 파종시기가 늦어져 영양생장 기간이 짧을수록 초장과 종실중이 감소하고(Lee et al., 2014a), 개화소요일수가 15일 이상 단축된다는 보고(Lee et al., 2021)와 일치한다.
작물 수량과 연관되는 형질인 종자수는 배주수(r=0.926***)와 고도로 유의한 정의 상관관계를 보였고, 협장(r=0.492***), 결실률(r=0.467***) 및 수당협수(r=0.191*)와도 유의한 상관성을 보였는데, 이는 협장이 길고 결실률이 높을수록 배주의 발달 공간 확보가 용이하여 배주수가 증가되고 종자 형성이 원활해지기 때문인 것으로 판단된다. 반면, 천립중의 경우 분지수(r=-0.273**) 및 결실률(r=-0.320***)과 유의한 부의 상관관계를 보였는데, 전 세계에서 수집한 488점의 유채 유전자원으로 농업적 형질을 조사하여 천립중과 분지수는 유의한 부의 상관관계를 보인다는 보고와 일치하였다(Chen et al., 2014). 또한, 수확 지수(harvest index)는 수량성 연구에 사용되는 특성으로 공급원-흡수원 비율(sink-source ratio)의 적절한 균형이 수확 지수에 영향을 미친다고 하였는데(Donald and Hamblin, 1976; Gifford and Evans, 1981; Luo et al., 2015), 분지수가 많거나 결실률이 높을수록 종자 발달 및 성숙에 필요한 양분의 공급이 분산되어 공급원-흡수원 비율 불균형이 초래되기 때문에 종자의 무게가 감소되는 상관관계가 있는 것으로 사료된다
주요 지방산 상관관계 분석에서 식용으로써 유채유의 품질에 중요한 올레인산 함량은 팔미트산(r=0.681***), 스테아르산(r=0.342***) 및 리놀레산(r=0.748***)과 높은 정의 상관관계를 보인 반면, 에이코젠산(r=-0.868***), 그리고 특히 에루스산(r=-0.984***)과는 고도로 유의한 부의 상관관계를 보였다. 각 유전자원 고유의 지방산 조성 중 에루스산(C22:1) 함량이 높을수록 올레인산(C18:1) 함량은 낮고, 반대로 에루스산 함량이 낮을수록 올레인산 함량이 높은 양상을 확인하였다(Fig. 1A). 올레인산과 에루스산은 전체 상관분석 결과 중 가장 높은 부의 상관성을 보였는데(Fig. 1B), 올레인산은 에루스산 생합성의 전구체로 색소체에서 합성된 올레인산이 세포질에서 에이코젠산을 거쳐 에루스산으로 합성된다(Katavic et al., 2002; Sinha et al., 2007). 따라서, 에루스산 함량이 높으면 올레인산이 전구체로 에루스산 생합성에 활용되어 함량이 낮아지고, 올레인산 함량이 높으면 그 반대의 관계로써, 올레인산과 에루스산의 함량은 부의 상관성을 가진다는 보고와 일치하는 결과였다(Nath et al., 2009; Rathod and Solanki, 2020; Tian et al., 2018). 유채 유전자원의 주요 지방산 조성 간 상관성에서 리놀렌산을 제외한 나머지 지방산들은 서로 간에 밀접한 상관관계를 나타내는 것을 확인하였다.
Table 3.
Correlation coefficients among 18 quantitative traits in 129 Brassica napus genetic resources.
| PHy | MBLx | NBw | NPv | PLu | NSt | NOs | SSRr | TSWq | Palp | Steo | Olen | Linm | Lnnl | Eick | Eruj | Others | |
| DFz | 0.63*** | 0.18* | -0.04 | 0.16 | 0.14 | 0.10 | 0.12 | 0.002 | 0.20* | 0.20* | -0.02 | 0.11 | 0.11 | -0.16 | -0.10 | -0.11 | -0.02 |
| PH | 0.45*** | -0.11 | 0.45*** | 0.36*** | 0.15 | 0.25** | -0.21* | 0.39*** | 0.16 | -0.11 | 0.08 | 0.01 | -0.28** | -0.04 | -0.06 | -0.10 | |
| MBL | -0.04 | 0.40*** | 0.19* | -0.04 | 0.04 | -0.20* | 0.29** | -0.04 | -0.15 | -0.11 | -0.18* | -0.15 | 0.16 | 0.13 | 0.02 | ||
| NB | 0.15 | -0.15 | -0.08 | -0.07 | -0.07 | -0.27** | 0.09 | 0.29** | -0.12 | -0.09 | -0.10 | 0.08 | 0.14 | -0.02 | |||
| NP | 0.26** | 0.19* | 0.28** | -0.18* | 0.10 | 0.22* | 0.09 | 0.19* | 0.07 | -0.26** | -0.09 | -0.17 | -0.28** | ||||
| PL | 0.49*** | 0.58*** | -0.06 | 0.17 | 0.07 | -0.16 | 0.10 | -0.08 | -0.12 | -0.01 | -0.08 | -0.20* | |||||
| NS | 0.93*** | 0.47*** | -0.11 | 0.41*** | 0.17 | 0.39*** | 0.26** | -0.13 | -0.28** | -0.38*** | -0.43*** | ||||||
| NO | 0.11 | 0.002 | 0.40*** | 0.13 | 0.40*** | 0.21* | -0.21* | -0.27** | -0.38*** | -0.48*** | |||||||
| SSR | -0.32*** | 0.14 | 0.17 | 0.09 | 0.20* | 0.17 | -0.10 | -0.13 | 0.01 | ||||||||
| TSW | 0.07 | -0.28** | 0.15 | -0.01 | -0.12 | -0.15 | -0.11 | -0.11 | |||||||||
| Pal | 0.46*** | 0.68*** | 0.76*** | 0.04 | -0.62*** | -0.74*** | -0.67*** | ||||||||||
| Ste | 0.34*** | 0.30** | -0.17 | -0.19* | -0.36*** | -0.50*** | |||||||||||
| Ole | 0.75*** | -0.23** | -0.87*** | -0.98*** | -0.80*** | ||||||||||||
| Lin | 0.13 | -0.76*** | -0.82*** | -0.56*** | |||||||||||||
| Lnn | 0.09 | 0.12 | 0.25** | ||||||||||||||
| Eic | 0.83*** | 0.58*** | |||||||||||||||
| Eru | 0.79*** | ||||||||||||||||
| Oth |
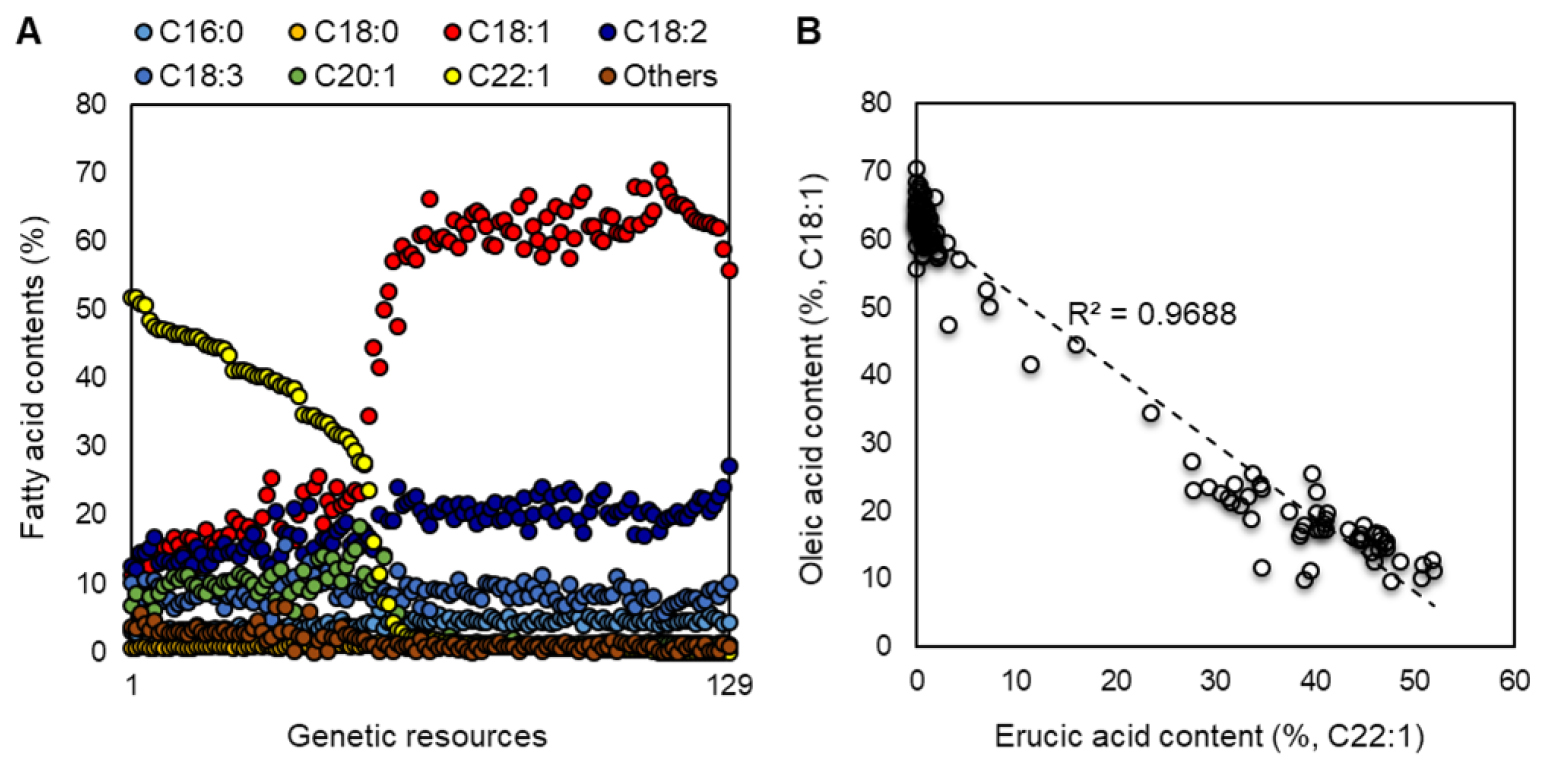
Fig. 1.
Fatty acids content analysis in 129 Brassica napus genetic resources. (A) Distribution of fatty acids in seeds. The x-axis represents the genetic resources in descending order of erucic acid content, while the y-axis depicts the percentage of fatty acid content. (B) Correlation analysis between oleic and erucic acids. A correlation coefficient was calculated using the Pearson’s correlation method.
주성분 분석
유채 유전자원 간의 유사관계를 분석하기 위해 10가지의 농업적 형질과 8종의 지방산 조성으로 주성분 분석으로 변수 간의 상호 의존도를 확인하였다(Table 4).
Table 4.
Eigenvalues and contribution obtained from principal component analysis in 129 Brassica napus germplasms.
고유값과 기여도를 볼 때, 제 1주성분은 전체 18 개 형질 중 5.6 개의 형질을 포함하며, 전체 변이의 31.29%를 설명할 수 있다. 제 2주성분은 2.9 개의 형질을 포함하며, 이는 제 2주성분까지의 누적 기여도로 전체 변이의 47.49%를 설명할 수 있다. 제 3주성분부터 제 5주성분까지 1 개 이상의 형질을 포함하며, 누적 기여도 73.90%의 전체 변이에 대한 설명력을 갖는 것을 확인하였다. 주성분 분석으로 차원 축소를 위해 최종 주성분의 개수를 선택함에 있어 변수의 변동량(variance)을 충분히 설명할 수 있는 누적 기여도(통상 80∼85% 이상)의 주성분 수를 선택하거나 주성분의 고유값이 1보다 큰 것에 해당하는 주성분들만을 선택하는 방법이 있다(Janmohammadi et al., 2014). 본 연구에서는 고유값이 1 이상을 기준으로 하여 제 5주성분까지 5 개의 주성분 수로 유전자원 특성 간 상관관계를 분석하였다(Table 5).
Table 5.
Five principal components among 18 quantitative traits in 129 Brassica napus germplasms.
| PC1 | PC2 | PC3 | PC4 | PC5 | |
| DFz(Days) | 0.085 | 0.278 | -0.115 | -0.022 | 0.619** |
| PHy(㎝) | 0.089 | 0.469** | -0.114 | 0.017 | 0.322 |
| MBLx(㎝) | -0.026 | 0.379* | -0.114 | 0.111 | 0.063 |
| NBw(no.) | -0.027 | -0.089 | 0.049 | 0.598** | 0.097 |
| NPv(no.) | 0.127 | 0.312 | -0.013 | 0.327 | 0.003 |
| PLu(㎝) | 0.102 | 0.341 | 0.322 | -0.161 | -0.207 |
| NSt(no.) | 0.256 | 0.101 | 0.525** | -0.099 | 0.021 |
| NOs(no.) | 0.256 | 0.203 | 0.437* | -0.049 | -0.140 |
| SSRr(%) | 0.073 | -0.217 | 0.370 | -0.159 | 0.424* |
| TSWq(g) | 0.044 | 0.292 | -0.355 | -0.263 | -0.170 |
| Palmitic (%) | 0.349 | -0.064 | -0.071 | 0.063 | 0.203 |
| Stearic (%) | 0.183 | -0.196 | 0.059 | 0.465* | 0.087 |
| Oleic (%) | 0.389* | -0.065 | -0.151 | -0.050 | -0.132 |
| Linoleic (%) | 0.331 | -0.190 | -0.173 | -0.118 | 0.174 |
| Linolenic (%) | -0.084 | -0.221 | 0.018 | -0.330 | 0.268 |
| Eicosenic (%) | -0.338 | 0.111 | 0.205 | 0.135 | 0.040 |
| Erucic (%) | -0.391** | 0.100 | 0.147 | 0.072 | 0.061 |
| Others (%) | -0.357 | 0.008 | 0.034 | -0.146 | 0.232 |
제 1주성분은 지방산 조성 요인에 높게 편중되어 있다. 올레인산, 팔미트산, 리놀레산과는 정의 상관관계를 보이고, 에루스산, 에이코젠산과는 부의 상관관계를 보였다. 특히 상호 간 상관성이 매우 높았던 올레인산과 에루스산의 함량이 가장 높은 연관을 보였다. 제 2주성분은 초장, 수장, 협장 등 길이 생장과 관련한 형질 요인과 정의 상관관계를 보였다. 제 3주성분은 꼬투리 및 종자의 발달 형질과 연관된 요인인 종자수 및 배주수와 정의 상관관계를 보이고, 천립중과는 부의 상관관계를 보였다. 제 4주성분은 발달적 측면의 형질인 분지수 및 수당협수, 그리고 스테아르산 함량과 정의 상관관계를 보였다. 제 5주성분은 개화소요일수 및 결실률 요인과 정의 상관관계를 보였다.
주성분 분석에서 기여도가 가장 높은 두 개의 주성분(PC1 및 PC2) 값에 대한 유채 유전자원의 분포를 2 차원 공간에 배열하여 시각화하였다(Fig. 2). 유전자원 변이의 31.3%를 설명하는 제 1주성분인 종자의 지방산 조성에 따라(x축) 좌측으로 에루스산 함량이 높은 유전자원들이 분포하고, 우측으로 올레인산 함량이 높은 유전자원들이 분포되었다. 에루스산과 올레인산은 서로 밀접한 부의 상관관계(R2=0.9688)가 있기 때문에 X축을 따라 좌우 양극단으로 나뉘어 분포되는 것을 확인하였다. 전체 변이의 16.2%를 설명하는 제 2주성분인 초장 및 수장 값에 따라(y축) 값이 큰 유전자원들은 상단에 분포하고, 값이 작은 유전자원들은 하단에 분포되었다. 주요 주성분에 기여하는 형질 및 개체별 분포로 유채 유전자원이 각각 어떤 변수에 영향을 많이 받는지 추정할 수 있다.
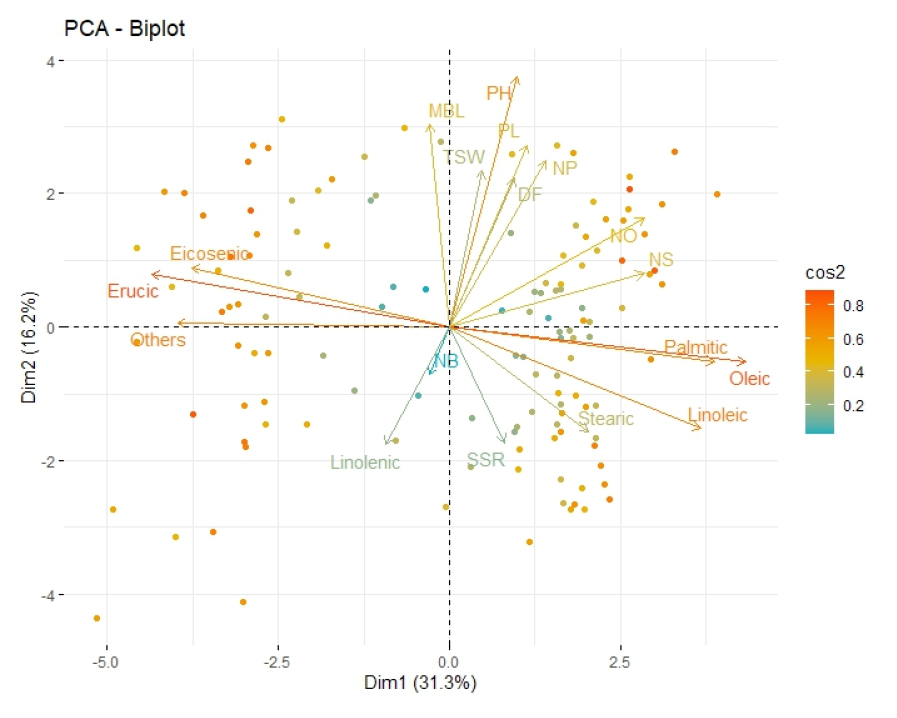
Fig. 2.
Principal component analysis (PCA) for agronomic traits and fatty acids of the seed in 129 Brassica napus resources. Biplot mixed on variables and individuals that contribute to the two-dimensional space. The colour including the length of arrow and the distance from zero point of dot are proportional to their contribution to the principal components.
군집분석
유채 129 개 유전자원의 10가지 주요 농업적 형질과 8종의 지방산 조성으로 수행한 주성분 분석 결과를 기반으로 제 1주성분에서 제 5주성분까지의 주성분 점수를 이용하여 군집분석을 수행하였다(Fig. 3). 군집 Ⅰ은 전체의 17.8%에 해당하는 23 자원이 속하고, 군집 Ⅱ는 전체의 28.7%를 차지하는 37 자원이 속하였다. 군집 Ⅲ은 전체의 31.0%에 해당하는 가장 큰 군집으로 40 자원이 이에 속하였다. 군집 Ⅳ는 전체 유전자원의 16.3%를 차지하는 21 자원이 속하고, 군집 Ⅴ는 전체의 6.2%를 차지하는 8 자원이 속한 가장 작은 군집이었다.
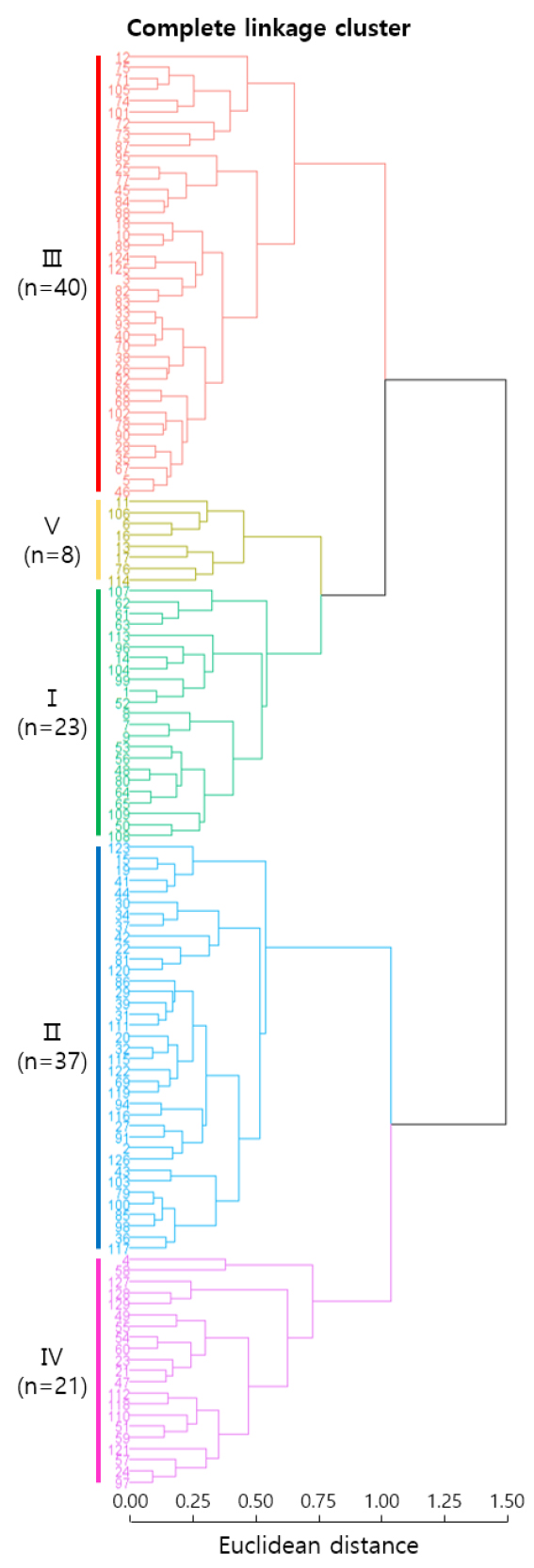
5 개 군집 간 양적 형질의 항목별 평균값의 결과는 Table 6과 같다. Ⅰ군집의 특징은 다른 군집보다 농업적 형질 중 수장이 가장 높고, 종자수는 가장 낮았으며, 지방산 조성 중 에루스산과 에이코젠산의 함량이 가장 높고, 스테아르산과 리놀레산이 가장 낮은 유전자원이 속한 군집이었다. Ⅱ군집은 종자수와 스테아르산 및 리놀레산 함량이 가장 높고, 에루스산과 기타 지방산 함량은 가장 낮은 군집이었다. Ⅲ군집은 협장과 배주수 및 천립중이 가장 높고, 분지수는 가장 낮았으며, 팔미트산과 올레인산 함량이 가장 높고, 에이코젠산 함량이 가장 낮았다. Ⅳ군집의 결실률 평균값은 가장 높은 반면, 농업적 형질 대부분의 값이 낮은 군집으로, 개화소요일수, 경장, 수장, 수당협수, 협장, 배주수 그리고 천립중이 다른 군집보다 낮았다. 지방산 조성은 리놀렌산과 기타 지방산 함량이 가장 높았고, 팔미트산과 올레인산 함량이 가장 낮은 자원이 속하였다. Ⅴ군집은 개화소요일수, 경장, 분지수 및 수당협수가 가장 높고, 결실률이 가장 낮았으며, 지방산 중 리놀렌산 함량이 가장 낮은 특성이 있었다.
Table 6.
Statistical mean value for agronomic traits and fatty acid contents of each cluster by complete linkage.
| Characters | Cluster (Mean) | ||||
| I | II | III | IV | V | |
| (n=23) | (n=37) | (n=40) | (n=21) | (n=8) | |
| DFz (Days) | 172.17 | 168.62 | 177.35 | 167.00 | 180.50 |
| PHy (㎝) | 149.00 | 131.87 | 163.18 | 124.86 | 182.42 |
| MBLx (㎝) | 56.45 | 44.27 | 53.78 | 43.76 | 54.00 |
| NBw (no.) | 9.26 | 9.85 | 8.47 | 9.63 | 12.96 |
| NPv (no.) | 64.68 | 56.21 | 68.20 | 46.11 | 70.33 |
| PLu (㎝) | 6.08 | 5.79 | 6.48 | 5.82 | 6.30 |
| NSt (no.) | 22.01 | 25.30 | 25.23 | 22.33 | 24.63 |
| NOs (no.) | 25.71 | 28.48 | 29.13 | 25.16 | 28.91 |
| SSRr (%) | 85.37 | 88.64 | 86.42 | 88.98 | 84.75 |
| TSWq (g) | 4.13 | 3.85 | 4.47 | 3.65 | 3.97 |
| Palmitic (%) | 3.53 | 4.52 | 4.64 | 3.50 | 3.89 |
| Stearic (%) | 0.84 | 1.05 | 0.96 | 0.85 | 0.92 |
| Oleic (%) | 18.17 | 60.99 | 61.65 | 17.23 | 19.28 |
| Linoleic (%) | 14.64 | 20.97 | 20.41 | 15.53 | 16.06 |
| Linolenic (%) | 8.57 | 8.51 | 8.40 | 9.86 | 7.63 |
| Eicosenic (%) | 10.81 | 2.06 | 1.80 | 9.85 | 9.31 |
| Erucic (%) | 40.63 | 1.22 | 1.38 | 40.06 | 40.39 |
| Others (%) | 2.80 | 0.68 | 0.76 | 3.14 | 2.52 |
우리나라 농업 상황에서 가장 적절한 유채 재배 양식은 논에서 이모작으로 가을 재배(10월 파종∼이듬해 6월 수확)하는 것이 기계화와 종자 생산량 측면에서 유리하다. 그러나, 가을 재배를 통한 논 이모작 작부체계는 논 벼와 재배기간 겹침 문제뿐만 아니라 최근 이상기상에 의한 가을철 유채 파종기에 잦은 강우와 큰 폭의 기온 일교차 등 발아 및 유묘 생육에 불리한 환경조건으로 적기에 유채 파종하기 어려운 경우가 많아 파종기가 뒤로 늦어지는 경우가 발생한다. 따라서, 현재 유채 품종 개발에서 우선되는 육종 목표는 개화소요일수의 단축이다. 또한, 유채 종자에서 추출한 유채유를 식품으로 활용하기 위해서는 에루스산의 함량이 2% 이하 수준으로 낮추는 것이 필수적이며, 저장, 유통, 품질 등의 이점으로 올레인산 함량이 높을수록 유리하다.
결과적으로, 군집 Ⅱ는 개화소요일수가 가장 짧은 군집 Ⅳ의 167일보다 1∼2일 정도밖에 차이 나지 않는 168.6일로 개화기가 빠른 집단이고, 에루스산의 함량이 1.22%로 전체 군집 중에서 가장 낮았으며, 올레인산 함량이 가장 높은 군집 Ⅲ의 61.65% 에 비해 0.66% 수준 차이에 불과한 60.99%의 매우 높은 고올레인산 군집이었다. 수량구성요소 중 하나인 종자수 또한 25.3 개로 가장 높은 군집이므로, 군집 Ⅱ의 37 자원을 활용한 우수 품종과의 교배 조합을 통해 품종 개발하는 것이 유채 농업 현황에 가장 유용할 것으로 판단된다.
적 요
본 연구는 유채 유전자원 129점에 대한 10가지 주요 농업적 형질과 8종의 종자 지방산 조성을 평가하고, 통계적 분석을 기반으로 자원을 군집화한 집단의 특징과 분포를 확인하였다. 이는 대량의 자원에서 통계적 신뢰성을 바탕으로 유의미한 특징을 도출하고, 군집화를 통해 육종에 필요한 형질을 가진 육종 소재를 1차적으로 집단 선발하는데 유용한 평가 방법이지만 일반적으로 작물은 동일한 장소에서 같은 품종을 재배하더라도 연차별 생육 특성이 온도, 강우, 병해충 등 환경조건에 따라 다소 차이를 보일 수 있다. 작물 재배의 생산 안정성은 작물의 형질과 환경조건과의 교호작용에 의해 품종마다 다르게 나타날 수 있어 통상적으로 연차간 균일한 생산력과 환경 적응성을 보이는 계통을 중심으로 품종을 육종한다. 따라서, 다량의 자원에서 유용 집단을 조속히 선별한 후 1차적으로 선발된 집단을 대상으로 연차간 생육 평가가 같이 병행되어야 정확한 육종이 수행될 것으로 판단된다. 이러한 결과는 추후 더 많은 유채 유전자원의 분석에 활용될 것이며, 육종 목표에 적합한 육종 소재 집단을 확보하는데 유용하게 이용될 것이다.
