

서 언
재료 및 방법
식물재료와 화기 구조 조사
웅예와 자예의 발달 특성
화분 활력 시기 조사
화서와 개체내 자가화합성 여부 조사
자가수정 혹은 교잡여부 확인을 위한 다형성 분자표지 개발
육종세대 단축 방법
결과 및 고찰
화서 발달 및 웅예선숙 특성
DAF 별 화기 발달 특성
화분의 활력과 임성 검증
단일화서 내 자가수정능력 검증
웅예선숙 극복을 위한 자가수정 시스템 확립
육종 세대 단축
적 요
서 언
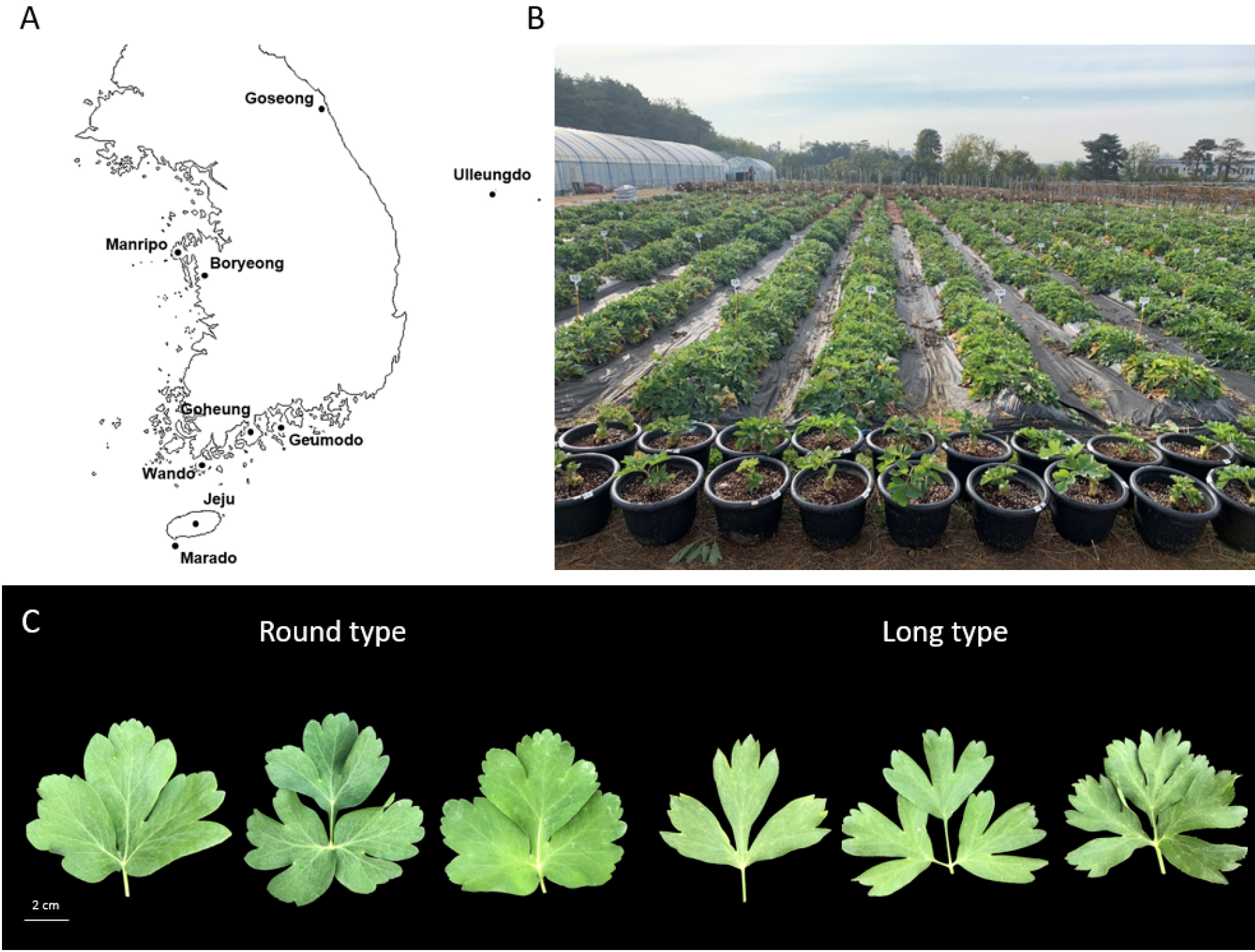
갯기름나물(Peucedanum japonicum Thunb.)은 산형과에 속하는 2년생식물로 한국, 필리핀, 일본, 중국 등에서 분포하며 한국에서는 중부 및 남부 해안 지역에 자생하고 있다(Lee et al., 2022a; Song and Cho, 2007; Yun et al., 2021). 갯기름나물의 국내 생산량은 2019년 기준으로 1,352 M/T이며, 재배면적은 137 ha 정도이다. 충남, 전북, 전남, 경북 내륙지역에서도 일부 재배되고 있으며, 전남 여수, 고흥, 금오도 등에서는 넓은 면적에 나물용으로 재배되고 있다.
갯기름나물의 잎과 줄기는 방풍나물이라고 부르기도 하며 독특한 맛과 향이 있어 산채 또는 나물로 이용되고 있다. <도문대작>에서는 방풍죽에 관해 극찬을 하였으며, <증보산림경제>에서 방풍 새싹으로 죽을 쑤면 그 맛이 매우 향미롭다고 나와 있다. 그 외에도 다수 옛 요리서에서 방풍요리에 대한 흔적이 발견되며 식용가치에 대해 오래전부터 인정되었다. 또한 뿌리는 한방에서 식방풍이라 불리며, peucedanol, umbelliferone등의 coumarin계 성분이 풍부하여 중풍, 감기, 신경통 등 질환을 치료하는 용도로 쓰이고 있다(Ikeshiro et al., 1992, 1993; Nam and Ryu, 1975; Shin et al., 2006). 일본에서는 장명초(長命草)라고 명명하며 다양한 건강기능성식품도 개발되어 이용되고 있다(長命草 <パウダー>N, Shiseido, Japan).
갯기름나물의 꽃은 복산형화서(Compound Umbel) 형태를 띄며 양성화이고 2~3년 주기로 개화하는 타식성작물로 비교적 긴 육종기간이 소요된다. 또한, 소화(Flower)는 개화와 더불어 웅예가 먼저 출현하여 성숙하고, 꽃잎과 웅예가 퇴화한 후에 자예가 출현하는 전형적인 웅예선숙(Protandry)의 개화특성을 가지고 있으며, 이러한 특성은 당근, 일당귀, 섬시호를 포함한 산형과 식물이나 국화과, 초롱꽃과 식물들에서도 나타난다(Lloyd and Yates, 1982; So et al., 2008; Sun and Ganders, 1988).
대부분의 약용식물은 주요 작물에 비해 유전 및 육종 연구가 거의 이루어지지 않고 있는 상태이다. 특히 타식성식물들은 집단 내 개체 간 변이가 많아 유전적 고정이 어려울 뿐만 아니라 육종기술도 미흡하며 품종 개발도 저조하다. 갯기름나물 역시 타식성식물로서 재배 과정에서 교잡이 이루어져 잎 모양, 줄기 색, 화서 형태(Umbel), 종자 형태 등에서 다양한 특성이 관찰된다. 농촌진흥청 경북농업기술원에서 1997년에 양질 다수성인 “식방풍 1호”를 육성하여, 품종등록하였지만(등록번호 제05-0026-1호), 이후 해당 품종의 보급 여부와 추가적인 품종 육성에 대한 정보는 부재하다(Chung, 1997). 여타의 나물들에 비해 칼슘과 칼륨 함량이 높다고 알려진 갯기름나물은 채소로서의 가치와 잠재력이 높기 때문에 고품질, 다수성, 고기능성의 우수한 특성을 지닌 갯기름나물의 품종 육성을 위해서는 이들 표현형에 대한 상세한 기준과 개화 습성을 정확히 파악하여 세대 촉진을 하기 위한 육종기술 개발이 필요하다(Jin et al., 2014; 2017). 본 연구는 잎의 향과 식감, 수량이 좋은 고품질 엽채소용 품종 혹은 기능 성분 함량과 뿌리 발육이 좋은 품종 등 채소와 약용 가치를 증진시킨 품종 등 특화된 우수 품종 육성의 기초자료로서 갯기름나물의 화기 구조, 개화 특성 및 웅예와 자예의 수정능력 등을 관찰하고, 이를 이용하여 효율적인 세대촉진 및 육종기술 개발을 도모하고자 하였다.
재료 및 방법
식물재료와 화기 구조 조사
고흥, 금오도, 완도, 제주도, 울릉도 등 각 지역에서 수집한 갯기름나물(식방풍) 개체들은 서울대학교 수원 농장(경기 수원시 권선구 탑동 540-478)과 서울대학교 수원 시험농장온실(경기 수원시 권선구 서호동로 13)에서 2년 이상 재배하며 개화 특성을 조사하였다(Fig. 1A, 1B). 갯기름나물 꽃에 대한 세부구조를 확인하기 위하여 온실에서 3~5월에 출현한 화서에서 웅예출현기부터 자예퇴화기까지 24시간 단위로 실체현미경(MD26-HD, Nikon, Japan)을 이용하여 웅예와 자예의 발달 특성을 관찰하였다. 웅예출현기는 웅예가 출현하고 성숙하여 화사의 신장, 약의 개열 후 퇴화되는 과정을 포함하는 시기이며, 이후 자예가 육안으로 확인되는 시기는 자예출현기, 출현한 자예가 성숙하는 시기를 자예성숙기, 주두의 끝이 구부러지며 활력이 거의 사라지는 시기를 자예퇴화기로 정의하였다(Chang et al., 1996b).
웅예와 자예의 발달 특성
개화기에는 웅예가 완전히 성숙하므로 웅예의 발달은 개화시를 통해 측정하였다. 개화 시기 및 특성을 구명하기 위해 같은 시기에 개화한 5개의 소화서(Umbellet)를 임의로 지정하여 웅예의 발달 단계를 관찰하였다. 첫 개화가 관찰된 소화서를 24시간 간격으로 관찰하며 개화한 꽃을 계수하였고, 개화율 및 개화패턴을 측정하였다.
개화시 자예는 전혀 발달하지 않은 상태이므로 자예의 성숙 정도는 자예의 인공수분 결과 종자의 결실율을 통해 확인하였다. 갯기름나물 식물체의 1차 화서를 대상으로 자예출현기 이후 1일 간격으로 한 개의 소화서를 대상으로 매일 인공수정을 하고 소화서 단위로 피봉을 하였다. 피봉전에는 자연방임이 가능하였고, 피봉 당일 외부 화분의 인공수정을 통해 수정능력을 극대화 하고자 하였다. 결실 후 약 30일 정도에 소화서간 결실된 종자들과 미결실 된 소화들을 계수하여 수분율과 주두의 수정능력을 추정하였다.
화분 활력 시기 조사
화분의 활력과 임성을 조사하기 위하여 기내에서 화분관 신장을 관찰하였다. 개화시의 화서 3개를 임의로 선정하여 약을 채취하였으며, 필터페이퍼가 깔린 페트리디쉬에 약을 올려놓고 뚜껑을 반쯤 연 상태로 25℃의 배양실에서 24시간동안 건조하여 개약하였다. 이후 뚜껑을 닫아 실온에 보관하였으며, 6일간 하루간격으로 화분의 활력을 측정하였다. 화분관 발아를 위해 증류수에 10% Sucrose, 200 g/L PEG (4000), 300 ㎎/L Boric acid, 300 ㎎/L Ca(No3)2, 200 ㎎/L MgSO4, 100 ㎎/L KNO3를 첨가한 후 pH 7로 적정한 것을 배양액으로 이용하였다(Wu and Gx, 2010). 개약 된 화분은 슬라이드 글라스 위에 도포한 후 배양액 20 ul를 위에 떨어뜨리고 커버글라스로 덮고 25℃의 항온 항습 조건에서 매 24시간 경과 후 화분관 발생을 조사하였다. 화분 발아 조사는 광학 현미경을 이용하여 80배율로 검경하였으며, 발아율은 1시야당 5~50립을 기준으로 5시야를 1반복으로 하고, 각 처리당 3반복으로 검사를 실시한 후 평균값을 백분율로 환산하였다. 화분 발아 판정은 화분관의 길이가 화분의 직경 이상으로 신장한 것을 기준으로 하였다(Cheon et al., 2006).
화서와 개체내 자가화합성 여부 조사
자가불화합성 식물로 알려진 갯기름나물의 자가화합가능여부를 관찰하기 위하여 2개의 계통을 임의로 선정하여 웅예출현기의 화서를 무제웅 상태에서 유산지격리하였으며, 자예퇴화기에 격리해제하여 30일 이후 결실여부를 조사하였다. 결실 된 종자가 자가수정 개체인지 확인하고자 결실된 종자는 트레이에 각 10립씩 파종하였으며, 발아된 5개체들의 잎으로부터 DNA를 추출하여 양친간 다형성을 보이는 5개의 분자마커를 적용하였다. 동일 화서뿐만 아니라 동일 개체 내에 존재하는 다른 화서 간 자가수정 여부를 확인하고 세대 촉진 방법을 적용하기 위하여 개체 내 주경과 지경 화서의 개화 시기를 조사 및 비교하였다. 조사는 정화서에서의 개화를 기점으로 이후 분지된 지경에 있는 화서들의 웅예출현기, 자예출현기와 자예퇴화기를 측정하였으며, 지경 간에 교배가능한 시기를 예측하였다.
자가수정 혹은 교잡여부 확인을 위한 다형성 분자표지 개발
다형성 분석을 위하여 갯기름나물 유전체 서열정보를 바탕으로 계통간 다형성을 보이는 5개의 dCAPS 분자표지를 제작하였다(Lee et al., 2022b; McKenna et al., 2010)(Table 1). 각 개체들에서 DNA를 추출한 뒤, 개발한 분자표지를 적용하여 자가수정 여부를 확인하였다. PCR 조건은 94℃에서 10분간 pre-denaturation시키고, 94℃에서 20초 denaturation, 59℃에서 20초 annealing 그리고 72℃에서 20초간 extension을 총 35회 반복한 뒤 마지막으로 72℃에서 7분간 extension을 진행하였다. 증폭된 PCR 산물은 각 마커 별로 적용되는 제한효소를 처리하여 37℃에서 over-night 한 뒤 최종 산물을 100 bp DNA ladder와 함께 3% agarose gel에서 100V로 50분간 전기영동하였다.
Table 1.
Marker information used for this study
육종세대 단축 방법
본 연구는 2018년도에 육성하여 수확한 84계통의 종자를 이용하였다. 관행방법은 2019년 1월 31일에 200공 트레이에 계통 당 50개씩 파종하였다. 생성된 묘들은 5월 1일에 계통 당 10개체씩 2반복으로 노지에 정식하고 생육하였으며, 노지에서 월동하고 이듬해 봄 개화와 종자성숙을 유도하였다. 세대 촉진을 위해 5월 15일에 200공 트레이에 파종하고 육묘 후 6월 18일에 직경 24 cm 포트에 정식하였으며, 10월 말까지 생장하고 1월 7일까지 자연 춘화처리가 되도록 하였다. 춘화처리후 20~25℃의 온도가 유지되는 온실로 포트를 이동하여 추대 및 개화를 유도하였다. 전체 84계통간 최초 형성된 정화서의 개화를 시작으로 온실 내 모든 개체들의 꽃이 개화할 때까지의 시기를 측정하였으며, 자가수정 및 인공교배를 진행하였다.
결과 및 고찰
화서 발달 및 웅예선숙 특성
갯기름나물은 주경 정단 및 각 가지의 끝에 화서가 형성된다. 한개의 화서는 약 20개의 소화서가 우산형으로 구성되어 있으며, 각 소화서에 존재하는 20~30개의 소화 또한 우산형으로 구성되어 있어 한 개의 화서에는 300~600개의 소화가 복산형화서 구조를 가진다(Fig. 2A). 일부 산형화과의 화서는 양성화와 수꽃이 공존하고 있다고 알려져 있지만 갯기름나물의 화서는 양성화만 존재하는 것으로 확인되었다(Doust, 1980; Hardin, 1929; Schlessman, 1976).
소화는 5개의 웅예와 2개의 자예가 존재하는 양성화이지만 꽃잎이 전개되는 것을 개화시로 정의할 때, 개화시에는 웅예만 발달하고, 자예는 미발달 상태로 관찰되지 않는다. 자예는 꽃잎과 웅예가 완전히 퇴화한 후 발달하는 전형적인 웅예선숙 특성을 보이며, 두개의 생식기관이 동시에 관찰되지 않는 자웅이숙 식물이다(Fig. 2B). 자웅이숙은 당귀나 시호와 같은 산형과 식물 대부분에서 관찰된다. 다른 웅예선숙 식물로는 수국과(Hydrangeaceae) 식물들이 존재하지만 이들은 웅예와 자예의 성숙시기는 틀리지만 두 기관을 동시에 관찰 할 수 있다(Choi, 2002).
소화서 내 소화들은 첫 개화 후 경과일수(Days After Flowering: DAF) 5일간에 5단계의 개화 패턴을 보였는데, 소화서의 가장 바깥쪽 소화로부터 1~5번 순서로 개화가 진행되었다(Fig. 2C, 2D). 그중 화서의 정중앙 소화(4번, Fig. 2D)는 대체로 5번보다 빨리 개화하지만 경우에 따라서는 3~5번 소화들이 동시에 개화하는 것이 관찰되었다. 산형과식물의 대부분은 바깥쪽 소화서가 중앙 소화서보다 결실 측면에서 우수하다고 알려져 있다(Webb, 1981). 섬바디나 일당귀는 갯기름나물과 유사하게 바깥쪽으로부터 안쪽으로 개화하는 특성을 보이지만 시호는 정중앙 소화의 개화를 시작으로 바깥쪽에서 안쪽으로 개화한다고 보고되고 있어 같은 산형과 식물 종 간에도 개화특성 차이가 존재함을 확인할 수 있다(Jung, 1993; Lee et al., 2020; Sun et al., 1997).
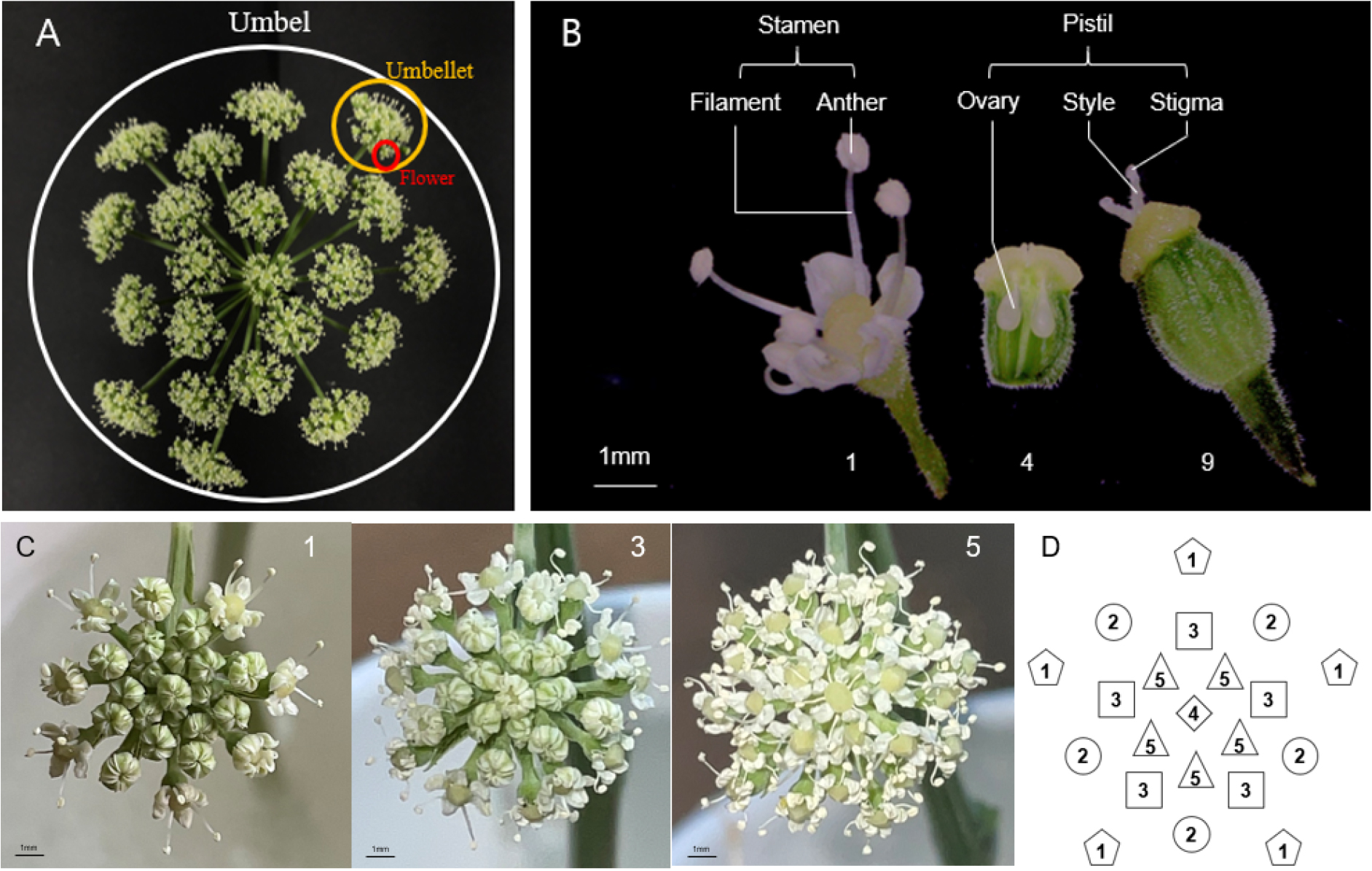
Fig. 2.
Flowering structure and process of P. japonicum. (A) Umbel structure of P. japonicum. White, orange and red circles indicate umbel, umbellet, and flower, respectively. Approximately 500~650 flowers in 15~25 umbellets are inspected in an umbel. (B) Male and female flower structure on different developmental stages. Numerical 1, 4, and 9 indicate days after flowering (DAF). DAF 4 flower was vertically cut to show ovary development. (C) Development of flowers in an umbellet at DAF 1, 3, and 5. (D) Schematic flowering order in an umbellet.
DAF 별 화기 발달 특성
갯기름나물은 바깥쪽 소화가 가장 먼저 개화하므로 소화서내 바깥쪽 소화가 첫 개화한 날을 DAF 1일로 설정하여 웅예의 발달과 자예의 성숙시기를 조사하여 적절한 수분시기를 확인하고자 하였다(Fig. 3). 소화서 내의 소화들은 DAF 1~5일동안 개화하며 화분의 성숙과 퇴화가 이루어지고, 자예는 DAF 8~13일 사이에 발달하여 임성을 띄기 시작하였다. 단일 소화서를 기준으로 DAF 1~5일차에 각각 20.1, 23.4, 23.4, 16.7, 16.4%의 비율로 소화가 개화하였으며, DAF 2~3일에 가장 왕성하였다(Fig. 3A).
웅예가 소멸하고 2~3일이 경과한 후 자예가 출현하였다. 자예가 출현하기 시작하는 DAF 7일부터 1일 간격으로 인공수분 후 피봉하여 각각의 DAF별 결실율을 조사하였다. 그 결과, DAF 8일부터 결실되었으며, 8~12일차 인공수분의 결실율은 각각 41, 69, 66, 56, 39%로 나타났다(Fig. 3C). DAF 9~11일의 결실률은 DAF 8일의 결실률보다 높은데, 이는 1~4일차에 개화한 소화들의 자예 임성이 해당 시기에 모두 유지되기 때문이라고 판단된다. DAF 10일 이후로부터 임성이 점차 감소하기 때문에 소화 1개의 자예는 개화 8일 후에 임성을 띄어 2일 정도 유지하고 퇴화하며, 개화 후 9~10일차 소화서의 임실율이 가장 양호할 것이라고 추정된다.
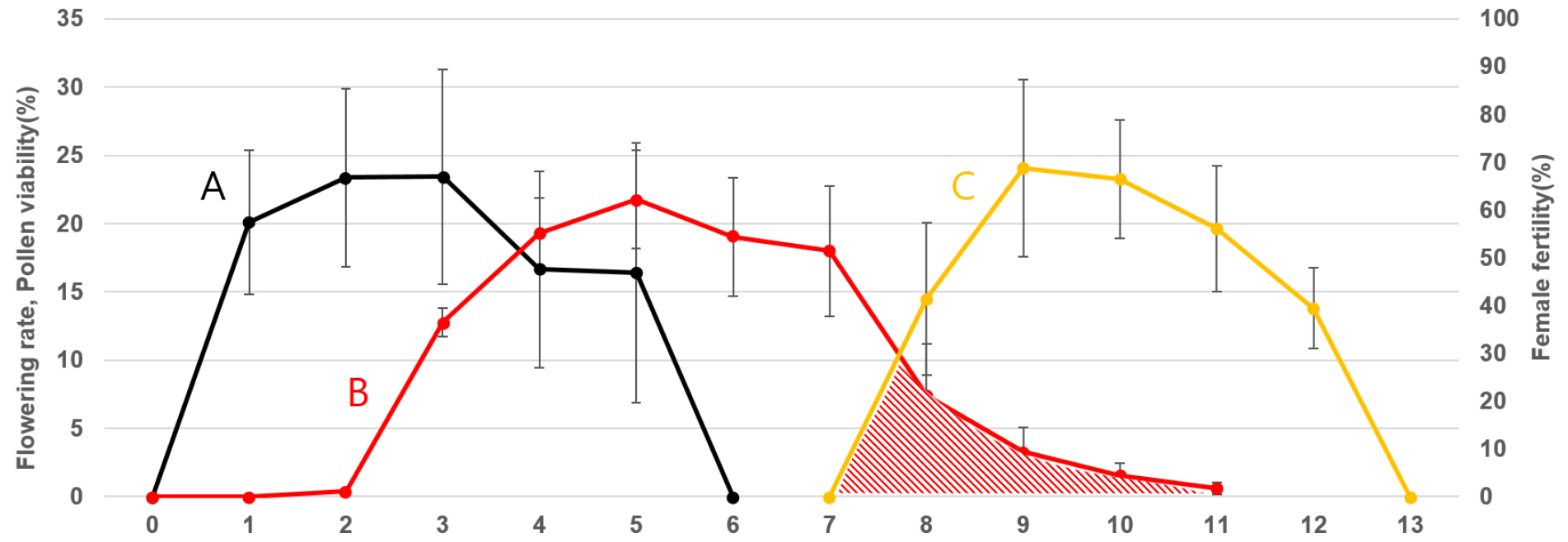
Fig. 3.
Development and fertility of male and female organs in a single umbellet. (A) The flowering rate in a umbellet for each DAF. (B) Pollen viability in a umbellet for each DAF. The pollen viability in a umbellet is calculated by reflecting the period required for the development of pollen and viability maintenance. (C) The fertility of pistil on each DAF. The overlapping period between B and C graph, denoted as shade, indicate the putative period for self-pollination.
화분의 활력과 임성 검증
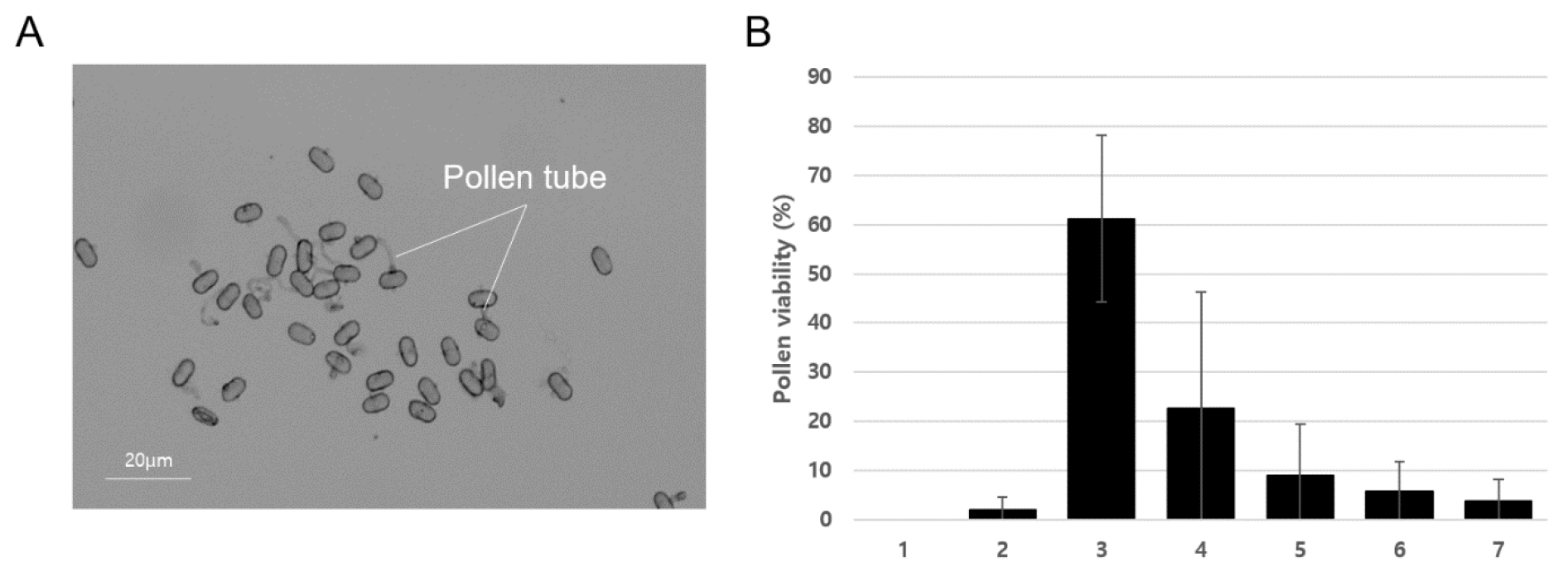
갯기름나물 화분의 임성 유지 기간을 알기 위하여 화분관의 활력이 얼마나 오래 유지되는지 측정하였다. 개화 직후 소화들로부터 채취한 화분들의 화분관 신장 측정을 통해 발아율을 확인하였다(Fig. 4). 화분채취 후 2~7일동안 각각 1.9, 61.3, 22.6, 9.0, 5.7, 3.7%의 발아율을 나타냈다. 이러한 결과로 보아 화분은 후숙 후에도 하루 정도의 성숙기간을 통해 임성이 증가하며 4~5일 정도 임성이 유지되는 것으로 추정된다.
단일 화서 내 화분의 활력은 화서 내 소화서의 발달기간과 화분의 활력 유지기간을 반영하여 추정할 수 있다. 이를 바탕으로 단일 화서 내 화분의 임성은 DAF 2~11일까지 유지가 될 수 있다고 판단된다(Fig. 3B). 반면, 화분의 임성유지기간과 주두의 가임기를 고려할 때 DAF 8~11일 사이 일부 단일 화서 내 자가 수정이 가능할 것으로 판단된다(Fig. 3).
단일화서 내 자가수정능력 검증
웅예선숙 식물의 소화는 자연 상태에서는 자예가 성숙기에 도달했을때 화분이 이미 비산하였거나 활력이 떨어진 상태가 되며, 이로 인해 자가수정 기회는 거의 없고 타가수정이 유도된다고 판단할 수 있다. 갯기름나물 이외에도 강활, 고본, 당귀, 백지, 시호, 천궁 등의 산형과 작물들은 타식성식물로 알려져있다(Ahn et al., 2011). 하지만 시호와 같은 일부 타식성식물들에게서 부분자식성도 확인되는 바, 갯기름나물의 화서 내 자가수정능력을 검증하고자 하였다(Chang et al., 1996a; Schlessman, 1981).
한 개의 화서를 개화시에 미리 피봉하여 외부로부터 화분의 유입을 차단하고 화서 내에서 생산된 웅예와 자예의 수정능력을 조사하였다. 계통마다 동일화서 내 임실율의 차이가 있었으나 0.4~4.4%의 자가수정 추정 종자가 결실되었다(Fig. 5A, 5B).
이후 계통간 다형성을 보이는 5종류의 DNA 마커를 이용하여 결실 종자가 자가수정 유래인지 확인하였다. 조사된 2계통의 자식 개체 5개체씩을 발아시킨 후 5개의 DNA 마커를 적용하였을 때 두 계통의 10개체 모두 자가수정시 나타날 수 있는 유전형을 보여주었다(Fig. 5C). 따라서 갯기름나물은 일반적으로 자연상태에서는 웅예선숙에 의한 자가불화합성이며 타식성이지만 부분적으로 자가수정도 가능한 부분자식성 식물로 판단할 수 있으며, 개체의 부위별 개화습성과 웅예선숙 특성을 이용한다면 개체 내 자가수정을 통한 고정계통 육성도 가능할 것으로 추정된다. 또한 결실된 종자들의 대부분이 소화서의 바깥쪽에 형성된 것을 확인 할 수 있었는데, 이는 앞서 제시한 DAF 4~5일차에 개화한 소화의 화분활력이 DAF 1~2일차에 개화했던 소화의 자예가 성숙할 때까지 유지되어 정상적인 수분이 되었다는 것을 의미한다(Fig. 5B). 따라서 화서 내 화분 활력기간과 자예의 성숙기간과 겹치는 8~11일 사이에 단일 화서 내 수분이 가능하다는 Fig. 3의 추정과 일치한다.
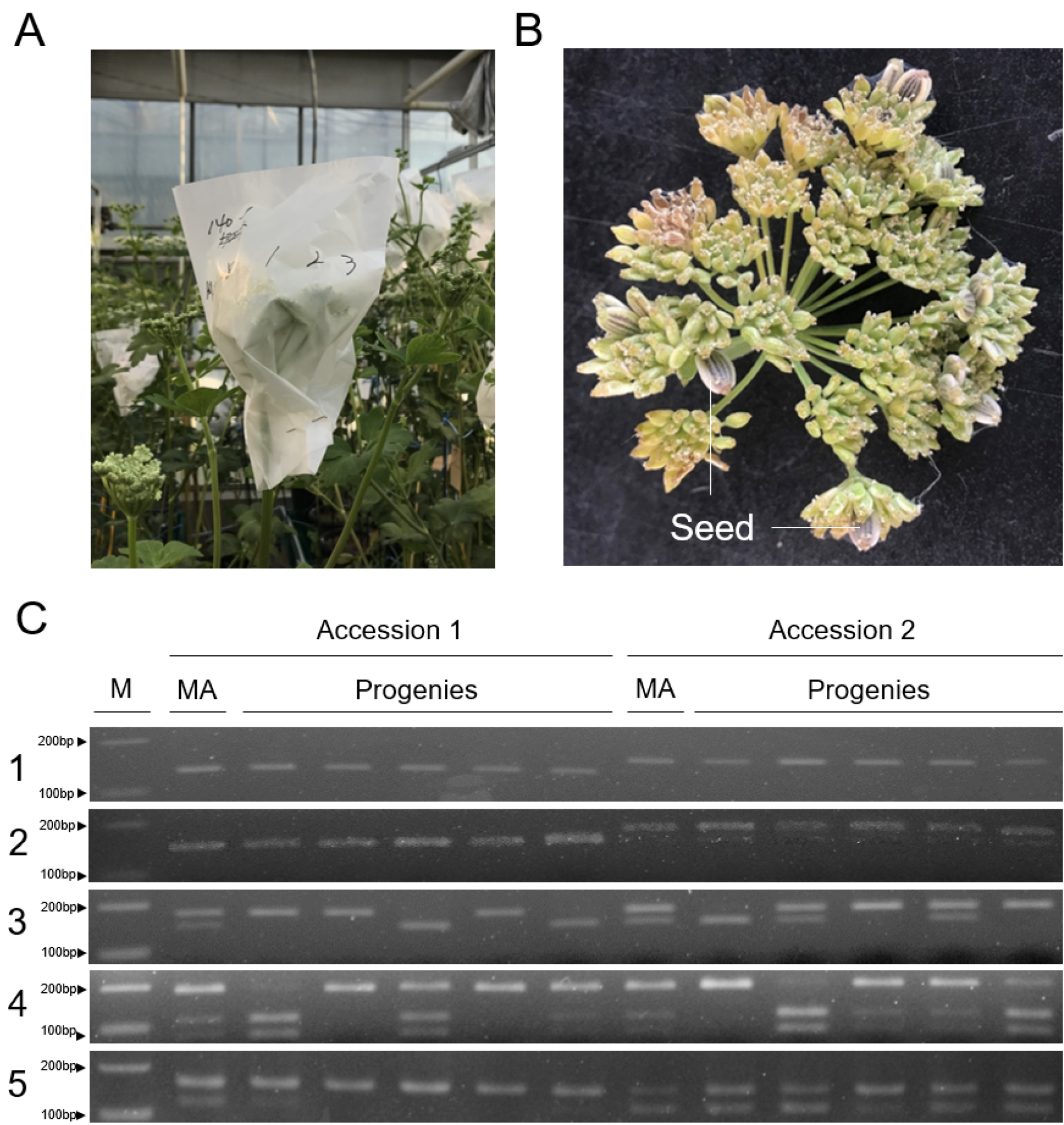
Fig. 5.
Induction and validation of self-pollination in an umbel. (A) Enveloping of an umbel for induction of self-pollination. (B) Seed sets in a solitary umbel. The seed formation rate of accession 1 and 2 is 0.42 and 4.41, respectively. (C) Validation of self-fertilization for progenies of seed sets from the solitary umbel. The number on the left represents the markers (Table 1). Five polymorphic molecular markers were applied for two different accessions. Markers 1 and 2 are homozygous in the maternal parent and showed identical genotypes among five progenies. Meanwhile, markers 3, 4, and 5 are heterozygous in the maternal parent and showed a segregation pattern among five progenies. M indicates a 100 bp DNA ladder. MA indicates genotypes of maternal accession. Progeny means each plant-derived seed formed in a solitary umbel.
웅예선숙 극복을 위한 자가수정 시스템 확립
위에서 확인된 갯기름나물의 부분자식성을 이용해 우수 계통육성을 위해 유전적 고정을 위한 자가수정 시스템을 확립하고자 하였다. 하지만 단일 화서 내에서 교배시 4.7% 이하의 낮은 자가수정률이 기대되므로 개체 내 다른 가지의 화서 간 자식 종자 확보가 가능하리라 판단된다(Dean et al., 1989; Webb, 1981).
갯기름나물의 한 개체는 주경의 끝에 정화서의 발생을 시작으로 계속된 분지를 통해 최대 4차 지경까지 화서가 형성되었으며, 정화서는 5일, 1·2차화서는 8일, 3차화서는 9일, 4차화서는 10일간 개화함으로써 지경이 많아질수록 긴 개화기간을 보였다. 이러한 개화기간의 연장으로 인해 자예의 임성 유지기간이 정화서는 8일, 1차화서는 10일, 2차화서는 12일, 3차화서는 13일, 4차화서는 10일간 지속되었다. 주경으로부터 분지된 1차화서는 정화서가 개화한 후 10일이 지나 개화하였으며, 2차 화서는 1차화서 개화 11일차에, 3차화서는 2차화서 개화 11일차에, 마지막으로 4차화서는 3차화서 개화 8일차에 개화가 시작되었다(Fig. 6).
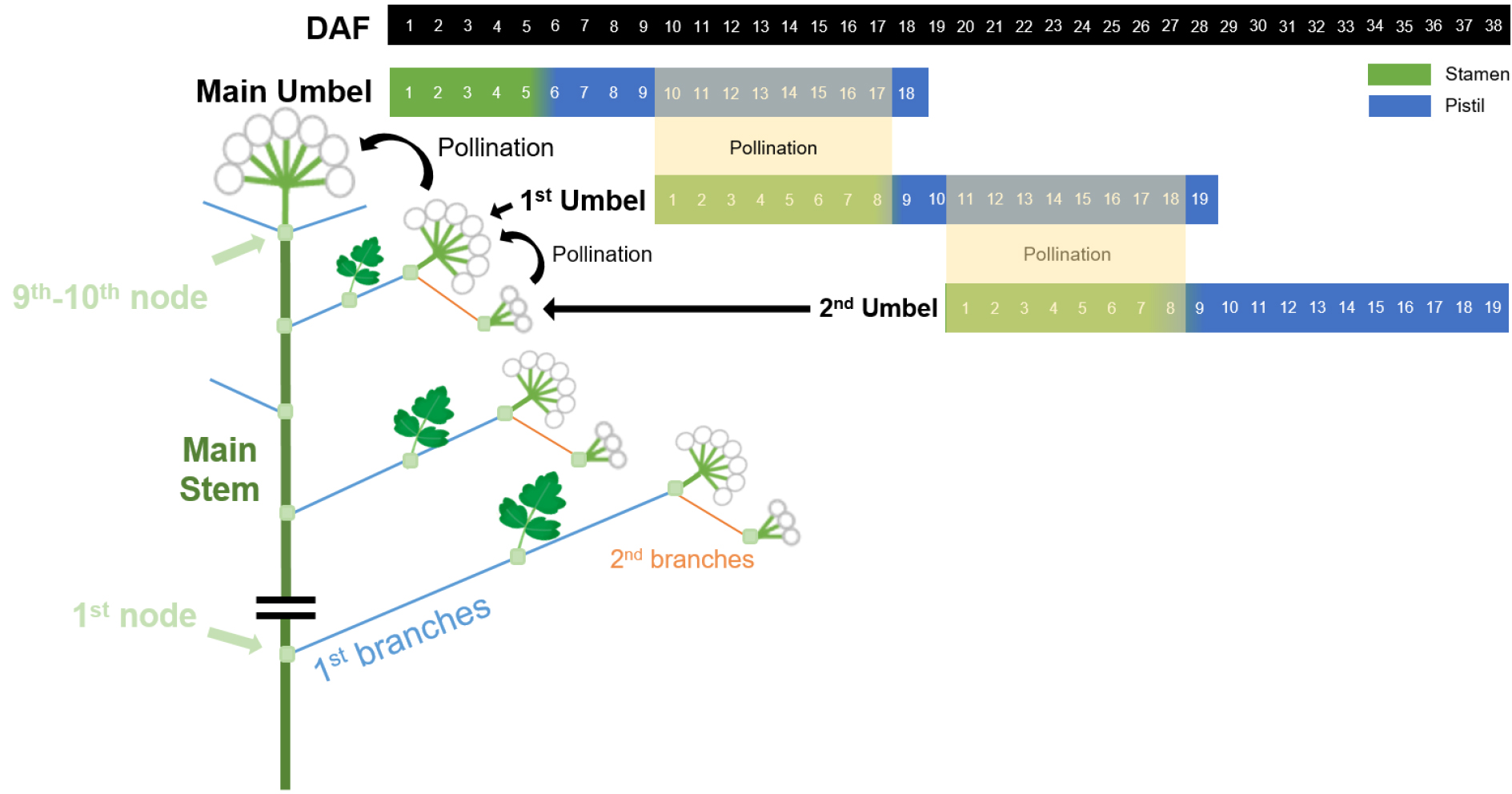
Fig. 6.
Flower development and fertilization characteristics on each branch of an individual P. japonicum plant. The branch umbels develop at the following branches at approximately 10~11 days intervals. Self-fertilization is possible by pollination between neighbor umbels during the periods of yellow box.
종합적으로 지경 화서 간 최소 8일의 개화시기 간격이 존재하였고 이로 인해 자예의 임성 기간과 화분의 활력 기간이 중복되므로 다른 가지의 화서 간 인공수정을 통해 자가수정 종자를 회득 할 수 있다. 정화서의 자예 활성과 1차 화서의 웅예출현기간은 9일간, 1차~2차 화서, 2차~3차 화서, 3차~4차 화서는 각각 9일, 9일, 12일간 중복되는 것으로 나타났다. 하지만 산형과 식물은 분지가 많아질수록 각 분지에 대한 영양 배분이 적어져 화서의 크기가 작아지고 종자의 임성도 저하되기 때문에 자식종자 채종을 위해서는 2차 지경 화서까지 이용하는 것이 적절할 것이다(Chang et al., 1996a; Koul et al., 1989; Lee et al., 2019).
육종 세대 단축
최근 밀, 유채 등 다양한 작물에서 육종세대를 비약적으로 단축할 수 있는 speed breeding 기술이 현실화되고 있으며, 특히 밀이나 보리의 경우 1년에 최대 6회 이상의 자가수정이 가능하다고 보고된 바 있다(Jähne et al., 2020; Samantara et al., 2022; Watson et al., 2018). 따라서 speed breeding 기술을 이용한다면 다양한 고정계통의 개발 및 교잡육종이 가능하므로 2년생 식물인 갯기름나물의 세대촉진 방법을 모색하였다.
갯기름나물의 노지재배는 영양생장 이후 10월 말부터 3월 초까지 약 20주간 월동을 통한 자연 춘화처리과정을 경과하게 되고 6월부터 개화가 시작되어 8~9월에 결실되어 채종한다. 노지 재배의 경우 월동에 의한 춘화처리 이후 점진적으로 상승하는 기온에 의해 개화기까지 많은 시간이 소요된다. 반면 온실재배의 경우 최소한의 저온을 경과한 후 온실에서 온도를 20~25℃로 유지시켜 줌으로써 신속한 영양생장과 생식생장을 촉진한다(Jin et al., 2020; Kim et al., 2019).
본 연구에서 이용된 84 육성계통은 완도, 금오도, 만리포, 고흥 등 남서해안 도서지역과 울릉도, 제주도 등 해안가 지역을 포함하여 전국에서 수집된 종자로부터 선발된 계통들로써 계통간 다양한 표현형 차이를 나타내었다(Fig. 1A, 1C). 잎의 모양은 둥근형~좁은타원형 타입이 관찰되었으며(Fig. 1C), 주경의 색깔은 초록색~자색, 흡지의 발생 정도가 많은 계통과 거의 없는 계통도 관찰되었다. 종자의 100립중도 0.35~0.97 g으로 다양한 차이를 보였다.
전체 84개의 육성계통들은 육묘 후 노지에서 정상 생육시킨 후 10월 중순에 각 계통별 한 개체씩을 화분으로 옮겨 심은 후 노지의 자연 저온을 통해 춘화처리과정을 거치고자 하였다(Fig. 1B). 산형과의 춘화처리는 일반적으로 11~12주의 기간이 필요로 한다는 선행연구에 따라 약 11주 정도 자연 저온 감응이 될 수 있도록 하였다(Atherton et al., 1990; Dias Tagliacozzo and Valio, 1994; Hiller and Kelly, 1979). 최근 3년의 수원지역 기상데이터에 따르면 10월 말부터 최저기온이 5℃이하로 떨어진다. 따라서 1월 초에는 약 11주 정도의 저온 감응 조건이 충족된다고 판단하여 1월 초에 84계통을 온실로 이동하여 개화를 유도하였다. 이를 통해 전체 84계통 중 74계통(88%)이 추대되었으며, 자가수정 혹은 인공교잡을 통한 종자채종이 가능하였다. 수집 자원 중 10계통은 저온 감응이 충분하지 않아 개화가 되지 않았는데, 이 자원들은 만추대 자원으로서의 가치가 있으므로 이듬해 노지에서 월동한 개체들로부터 종자채종을 하였다.
추대가 진행된 74개체는 온실 입실 기준으로 24~28일이 경과한 후 개화하였다. 가장 빨리 개화한 개체는 온실 입실 기준으로 9일이 경과된 1월 15일에 최초로 화뢰가 형성되었고, 약 15일 후인 1월 30일에 정화서가 개화하였다. 대부분의 개체는 4월까지 2달간 자가수정 혹은 인공교잡이 가능하였고 5월에 종자가 성숙되어 채종이 가능하였다. 채종된 종자는 정선을 거친 후 5~6월에 재파종하여 이듬해 5월에 다시 종자를 확보할 수 있어 1년 1세대 진전이 가능하였으며, 관행의 방법에서 2년이 소요되던 세대기간을 반으로 단축할 수 있는 육종시스템을 확립하였다(Fig. 7).
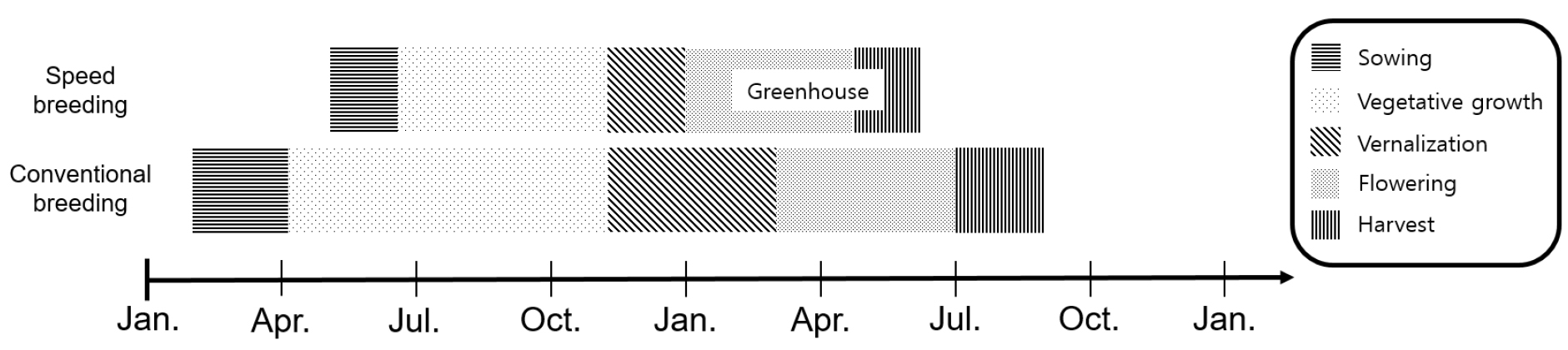
Fig. 7.
Graph depicting the development stages of speed breeding and conventional breeding. Speed breeding promotes flowering by transferring the cold acclimated plants into the greenhouse. The plants flower on January and the seeds can be harvested on May. The seeds can be immediately propagated for next generation, reducing the breeding cycle from two years to one year.
적 요
본 연구는 갯기름나물의 개화특성 및 수정능력을 파악하여 육종의 기초자료로 이용하고자 수행되었다. 개화기 관찰 결과, 소화서 내 소화들은 바깥쪽에서부터 중앙쪽 방향으로 5일의 시차를 두고 개화하였다. 단일 소화는 웅예선숙형 특성을 가지며 개화와 더불어 화분은 5일간 유지되었고, 자예는 개화후 8일차부터 발달하여 13일까지 임성을 유지하였다. 화분은 웅예출현 직후 2일째에 활력이 대폭 증가하였지만 하루만에 급감하였다. 동일화서 내 자가수정능력을 확인한 결과, 일부에서 자식성종자를 결실하였고, DNA 마커에 의해 자가수정 개체임이 확인되었다. 개체내 분지는 주경을 기점으로 영양상태에 따라 최대 4차지경까지 분지되며, 지경 화서 간 평균 8일 동안 자예 활성, 화분 활력시기가 중복되어 다른 가지의 화서간 자가수정이 가능하다고 판단된다. 노지에서 생육 후 12월 말까지 춘화처리 된 개체들을 12월 말에 온실내에서 생식생장을 유도하고 5월에 채종함으로써 개화시기를 약 4개월정도 단축시킬 수 있었다. 채종한 종자는 5~6월에 파종하고 영양생장 후 12월 말까지 춘화처리 후 5월에 채종할 수 있어 기존 2년 1세대 육종기간을 1년 1세대로 단축할 수 있었다.
