

서 언
재료 및 방법
실험재료
시료추출
세포배양
세포독성 측정
Nitric oxide (NO) 측정
포식작용(Phagocytosis) 측정
SDS-PAGE and Western blot analysis
Reverse transcriptase-polymerase chain reaction (RT-PCR)
통계분석
결과 및 고찰
인가목 잎 추출물의 대식세포 활성화 유도
인가목 잎 추출물의 대식세포 자가포식 유도
Toll-like receptor 4 (TLR4)의 인가목 잎 추출물에 의한 대식세포의 자가포식 유도에 미치는 영향
PI3K/AKT 신호전달의 인가목 잎 추출물에 의한 대식세포의 자가포식 유도에 미치는 영향
적 요
서 언
인간은 내·외부에 존재하는 다양한 미생물과 공존하며 살아가고 있다. 그럼에도 불구하고 인간은 선천적 및 적응적 면역 체계를 가지고 있어서 병원체로부터 야기되는 다양한 질병을 회피한다(Hirayama et al., 2018). 인체의 선천성 면역 반응은 감염초기에 병원체들을 인식하고 제거하는 역할을 하며, 적응성 면역 반응은 항원 특이적 항체를 생성하여 병원체들을 제거한다(Hirayama et al., 2018). 그러므로 선천성 및 적응성 면역반응의 활성화는 신체의 항상성 유지에 필수적이라고 알려져 있다. 선천성 면역 반응에 관여하는 세포인 대식세포는 다양한 막 수용체를 통해 외래 병원체를 인식하고, 식균작용을 통해 침입한 외래 병원체를 제거한다(Schenten and Medzhitov, 2011). 또한, 대식세포가 분비하는 전 염증성 사이토카인들은 적응성 면역반응에 관여하는 T 세포 및 B 세포의 활성화를 유도한다고 알려져 있다(Duque and Descoteaux, 2014). 그리고 대식세포는 적응성 면역세포에 병원체의 정보를 제공하는 항원제시기능을 가지고 있다(Barker et al., 2002). 따라서 대식세포의 활성화는 선천면역과 적응면역반응을 동시에 증가시키는 좋은 전략이라고 여겨지고 있다.
자가포식(Autophagy)은 분해를 통한 재활용 과정으로 생명체의 정상적인 생물학적 기능을 유지하는 데 필수적이라고 알려져 있다(Wu and Lu, 2020). 대식세포의 자가포식은 인체가 다양한 병원체에 대항하는 주요 면역기전 중 하나로 알려져 있고, 대식세포의 주요 기능인 식균작용, 전 염증성 사이토카인 방출, 염증 반응 및 항원 제시 기능에 관여한다(Wu and Lu, 2020). 게다가 대식세포에서 자가포식 유도는 대식세포의 항원 처리 과정 및 항원 제시기능을 강화해 선천성 면역 반응과 적응성 면역 반응 사이의 연결을 증가시킨다고 알려져 있다(Kirkegaard et al., 2004). 대식세포의 자가포식 유도는 적응성 면역 세포의 항체 생성에 관여하기 때문에, 대식세포의 자가포식을 촉진 시키는 물질들은 면역 반응을 강화하기 위한 잠재적인 백신 보조제로 활용될 수 있다고 보고되고 있다(Bah and Vergne, 2017).
인가목(Rosa acicularis Lindl.)은 장미과(Rosaceae)에 속하는 낙엽성 관목으로 중국, 일본, 러시아, 몽고, 카자흐스탄 북유럽, 북아메리카 및 한국 등지에서 자생하고 있으며, 인가목의 잎, 꽃, 뿌리 및 열매는 식품과 약용식물로 활용되어왔다(Olennikov et al., 2021). 인가목의 열매는 잇몸 강화와 심장 치료, 인가목의 가지는 설사와 장 질환 예방 그리고 인가목의 줄기 껍질은 해독제 및 림프계 질환 치료를 위해 사용되어 왔다(Olennikov et al., 2021). 최근 연구를 통해 인가목은 항산화 활성, 지질분해효소 활성억제 및 항균활성이 있다고 보고되고 있다(Olennikov et al., 2021). 그러나 인가목의 면역증진활성 관련 연구는 전무한 실정이다. COVID-19가 성행하면서 면역증진에 관한 관심이 높고 면역증진 소재의 발굴이 중요시되고 있으므로 본 연구에서는 인가목의 대식세포 활성화에 대한 영향을 평가하였다.
재료 및 방법
실험재료
MTT, Griess 및 화학적 억제제(TAK-242, C29, PD98059, SB203580, SP600125, BAY 11-7082 및 LY294002)는 Sigma-Aldrich (St. Louis, MO, USA)에서 구입하였다. 1차 항체(p-JNK, p-PI3K, p-AKT, p62/SQSTM1 및 β-액틴) 그리고 2차 항체(Anti-rabbit)는 Cell Signaling (Bervely, MA, USA)에서 구입하였다.
시료추출
인가목(Rosa acicularis Lindl.) 잎은 국립산림과학원 산림약용자원연구소에서 식물학적 동정 후 제공받았다. 건조 후 파쇄된 인가목 잎을 부피의 20배증류수로 40℃에서 24시간 동안 추출하였다. 추출 후 원심분리하여 획득한 추출물을 동결 건조하였다. 동결건조 된 인가목 잎 추출물 (추출수율: 22.45%, RAL)은 증류수로 용해하여 실험에 사용하였다.
세포배양
American Type Culture Collection (Manassas, VA, USA)에서 구입한 마우스 대식세포인 RAW264.7 세포는 10% FBS, 100 U/mL penicillin과 100 ㎍/mL streptomycin이 포함된 DMEM/F-12 배지로 37℃, 5% CO2 하에서 배양되었다.
세포독성 측정
인가목 잎 추출물의 RAW264.7세포에 대한 세포독성은 MTT assay로 측정하였다(Geum and Jeong, 2021). RAW264.7세포를 96-well plate에 배양하여 세포가 well에 80%이상 찼을때, 인가목 잎 추출물을 농도별로 24시간 동안 처리하였다. 인가목 잎 추출물 처리 24시간 후, 각 well에 MTT 용액(1 ㎎/mL)을 각 well에 50 μL를 첨가하고 4시간동안 반응시켰다. 4시간 후 상등액을 제거하고 각 well에 DMSO를 100 μL씩 첨가하여 세포에 흡수된 MTT를 용출시켜 UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 570 ㎚에서 흡광도를 측정하였다.
Nitric oxide (NO) 측정
인가목 잎 추출물의 RAW264.7가 분비하는 NO 생성에 미치는 영향은 Griess assay로 측정하였다(Geum and Jeong, 2021). RAW264.7세포를 12-well plate에 배양하여 세포가 well에 70%이상 찼을때, 인가목 잎 추출물을 농도별로 24시간 동안 처리하였다. 인가목 잎 추출물 처리 24시간 후, 세포배양액과 Griess 시약(Sigma-Aldrich, St. Louis, MO, USA)을 1:1 비율로 희석하여 상온에서 15분간 반응시킨 후, UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 540 ㎚에서 흡광도를 측정하였다.
포식작용(Phagocytosis) 측정
인가목 잎 추출물의 RAW264.7세포의 포식작용에 대한 영향은 neutral red assay로 측정하였다(Geum and Jeong, 2021). RAW264.7세포를 12-well plate에 배양하여 세포가 well에 70%이상 찼을 때, 인가목 잎 추출물을 농도별로 24시간 동안 처리하였다. 인가목 잎 추출물 처리 24시간 후, 세포배양액을 제거하고 500 μL의 0.1% neutral red 용액(Sigma-Aldrich, St. Louis, MO, USA)으로 37℃에서 3시간 동안 세포를 염색시켰다. 3시간 후, 각 well에 lysis buffer (ethanol/acetic acid = 1:1) 500 μL를 첨가하여 세포로 흡수된 neutral red를 용출시켜 UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 540 ㎚에서 흡광도를 측정하였다.
SDS-PAGE and Western blot analysis
RAW264.7 세포를 1 × phosphate-buffered saline으로 2회 세척한 후, protease inhibitor cocktail (Sigma-Aldrich Co.)과 phosphatase inhibitor cocktail (Sigma-Aldrich Co.)이 포함된 radioimmunoprecipitation buffer (Boston Bio Products, Ashland, MA, USA)로 RAW264.7 세포로부터 단백질을 추출하였다. 추출된 단백질은 Bicinchoninic acid protein assay (Pierce Biotechnology Inc., Waltham, MA, USA)로 정량분석하였고, 동일량의 단백질을 SDS-acrylamide gel로 전기영동하고 PVDF membrane (Bio-Rad, Hercules, CA, USA)에 이동시킨 후 5% non-fat dry milk로 상온에서 1시간 동안 blocking 하였다. 1시간 후, 1차 항체를 5% non-fat dry milk에 용해시켜 4℃에서 16시간 동안 반응시킨 후 membrane을 0.05% tween-20이 포함된 tris-buffered saline (TBS-T)로 5분간 3회 세척하였다. 그 후 2차 항체는 5% non-fat dry milk에 용해시켜 membrane에 상온에서 1시간 처리하였고, TBS-T로 5분간 3회 세척 후 membrane은 ECL western blotting substrate (Amersham Biosciences Co., Little Chalfont, England)를 이용하여 단백질을 확인하였다.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
RAW264.7 세포를 1 × PBS로 2회 세척한 후, RNeasy Mini kit (QIAGEN GmbH., Hilden, Germany)를 이용하여 RAW264.7 세포로부터 total RNA를 추출하였다. 1 ㎍의 total RNA를 Verso cDNA synthesis kit (Thermo Fisher Scientific Inc., Waltham, MA, USA)를 이용하여 cDNA를 합성하였다. PCR은 PCR Master Mix Kit (Promega Co., Madison, WI, USA)를 이용하여 수행되었고, 사용된 primer는 Table 1과 같다.
Table 1.
Sequence of oligonucleotide primers used for RT-PCR
통계분석
모든 결과는 3회 반복 측정 후 평균 ± 표준편차로 나타내었고, 처리간 유의성은 Student’s t-test로 검증하여 p-value 값이 0.05 미만일 때 통계적으로 유의하다고 판정하였다(Microsoft Exel 2010, Microsoft, Redmond, WA, USA).
결과 및 고찰
인가목 잎 추출물의 대식세포 활성화 유도
인체의 면역체계는 감염 초기에 외래 병원체를 인식하고 제거하는 선천성 면역체계과 세포독성 반응과 항원 특이 항체를 분비하여 외래 병원체를 제거하는 적응성 면역체계로 구성되어 있다(Hirayama et al., 2018). 선천성 면역세포 중 대식세포는 외래 병원체 침입 시 활성화되어 면역자극인자의 분비와 포식작용을 통해 외래 병원체들을 제거한다고 알려져 있다(Sieweke and Allen, 2013). 그러므로 대식세포의 활성화는 인체의 선천성 면역체계의 강화와 밀접한 연관이 있다고 알려져 있다.
인가목 잎 추출물(RAL)이 대식세포의 활성화를 유도하는지 확인하기 위해 RAW264.7세포에 인가목 잎 추출물을 처리한 후 RAW264.7세포가 분비하는 NO, iNOS 그리고 IL-1β의 생성양과 대표적인 대식세포 활성화 지표인 포식작용을 분석하였다. 그 결과, 인가목 잎 추출물은 RAW264.7 세포에서 외래 병원체를 파괴하는 NO와 iNOS 그리고 다양한 선천성 면역세포와 적응성 면역세포의 활성화를 유도하는 IL-1β의 생성을 증가시켰다(Fig. 1A). 그리고 인가목 잎 추출물이 처리된 RAW264.7 세포에서 포식작용의 활성화가 유도되었다(Fig. 1B). 대식세포가 분비하는 면역자극인자들은 신체의 면역체게 강화와 밀접한 관련이 있으나 과도한 면역자극인자들의 분비는 세포독성을 유발한다고 알려져 있다(Geum and Jeong, 2021). 그리하여 인가목 잎 추출물의 RAW264.7 세포에 대한 세포독성을 확인한 결과, 인가목 잎 추출물은 RAW264.7 세포에 대한 세포독성이 없는 것으로 나타났다(Fig. 1C). 본 결과를 미루어 볼 때, 인가목 잎 추출물은 인체의 면역시스템 강화를 위한 기능성 식의약 소재로 활용이 가능할 것으로 판단된다.
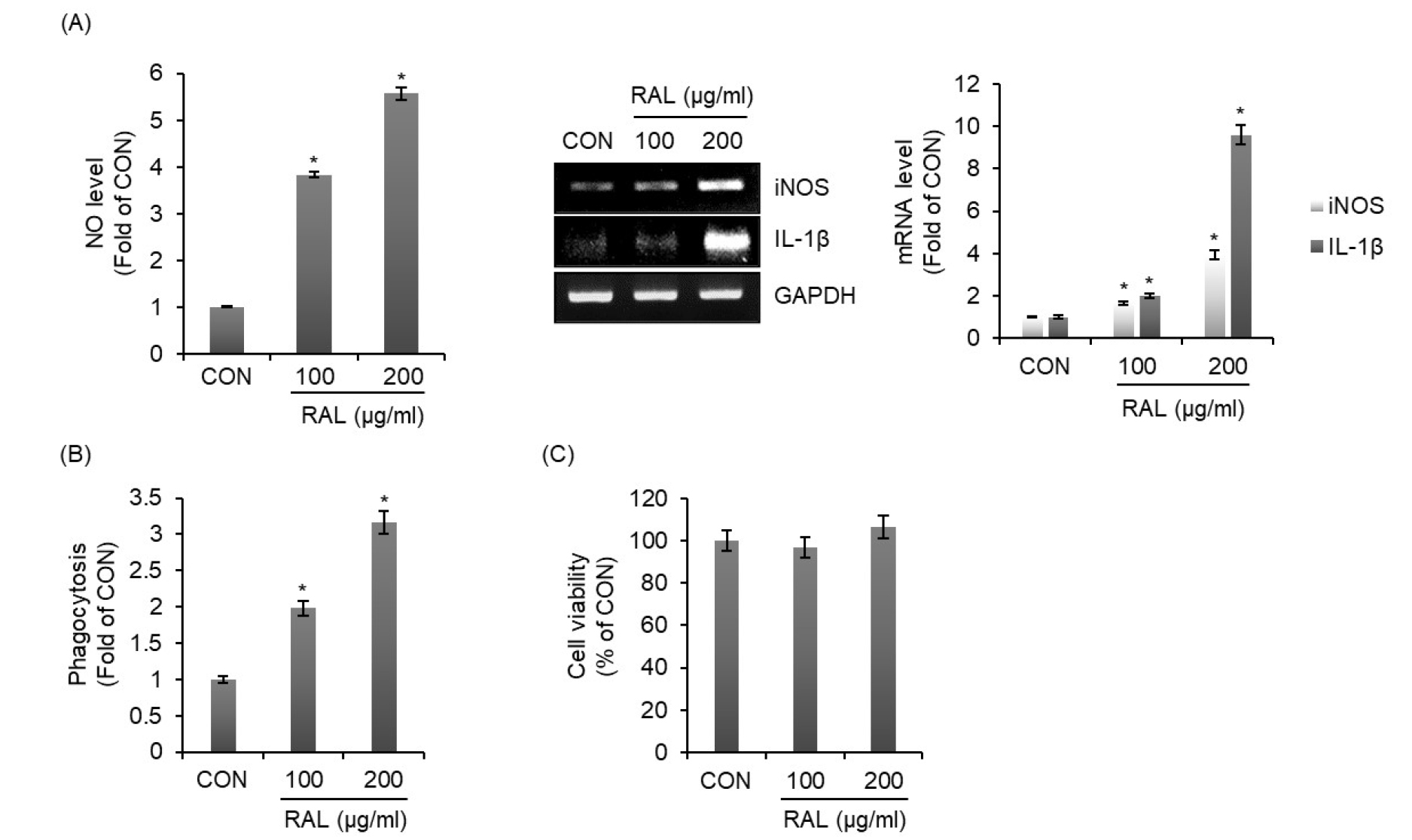
인가목 잎 추출물의 대식세포 자가포식 유도
외래 병원체에 대한 활성화된 대식세포의 주요기능인 외래 병원체를 인식하는 항원 인식 능, 외래 병원체의 정보를 항체를 생성하는 적응성 면역세포에 전달하는 항원 제시기능 그리고 외래 병원체를 제거하는 포식작용과 면역자극인자들의 분비기능들은 대식세포의 자가포식과 밀접한 관련이 있다고 보고되고 있다(Wu and Lu, 2020). 대식세포의 자가포식은 백신 개발 측면에서도 중요하게 생각되고 있다(Chou et al., 2020). 백신에는 외래 병원체인 항원과 항원 특이적 항체반응을 유도하는 보조제가 포함되어 있는데, 보조제는 T 세포와 B 세포의 항원 특이적 적응성 면역 반응을 활성화하기 위해 필수적으로 요구된다(Chou et al., 2020). 대식세포의 자가포식 유도를 통한 항원 제시기능 강화와 T 세포의 분화촉진은 초기 면역 반응을 유도하는 백신 보조제 개발의 주요 전략으로 활용되고 있다(Coffman et al., 2010). p62/sequestosome 1 (p62/SQSTM1)은 자가포식 과정에서 중요한 cargo receptor로 cargo를 유비퀴틴화된 응집체로 전환시켜 자가포식소체에 격리하는 역할을 한다(Komatsu and Ichimura, 2010; Lamark et al., 2009). p62/SQSTM1은 침입하는 외래 병원체를 표적하기 때문에 자가포식에 중요하다고 알려져 있다(Zheng et al., 2009). 그리하여 p62/SQSTM1은 자가포식 활성의 주요 지표로 활용되고 있다(Liu et al., 2016).
인가목 잎 추출물이 대식세포 자가포식에 미치는 영향을 조사하기 위해 인가목 잎 추출물이 처리된 RAW264.7 세포에서 p62/SQSTM1 단백질의 수준을 측정하였다. 그 결과, 인가목 잎 추출물은 처리 3시간부터 p62/SQSTM1의 단백질 수준을 증가시켰다(Fig. 2A). 그리고 인가목 잎 추출물은 농도 의존적으로 p62/SQSTM1의 단백질 수준을 증가시켰다(Fig. 2B). 본 결과를 미루어 볼 때, 인가목 잎 추출물은 대식세포의 자가포식 유도에 긍정적인 영향을 미칠 수 있음을 보여준다. 최근 rapamycin은 대식세포에서 자가포식을 유도한다고 보고되고 있어 백신 보조제 활용을 위한 임상적 연구가 진행되고 있다(Huang et al., 2017). 그러나 rapamycin은 다양한 부작용을 초래해 백신보조제 활용에 대한 제한점이 있다. 그리하여 인가목 잎 추출물은 부작용이 없는 백신보조제로 활용이 가능할 것으로 판단된다.
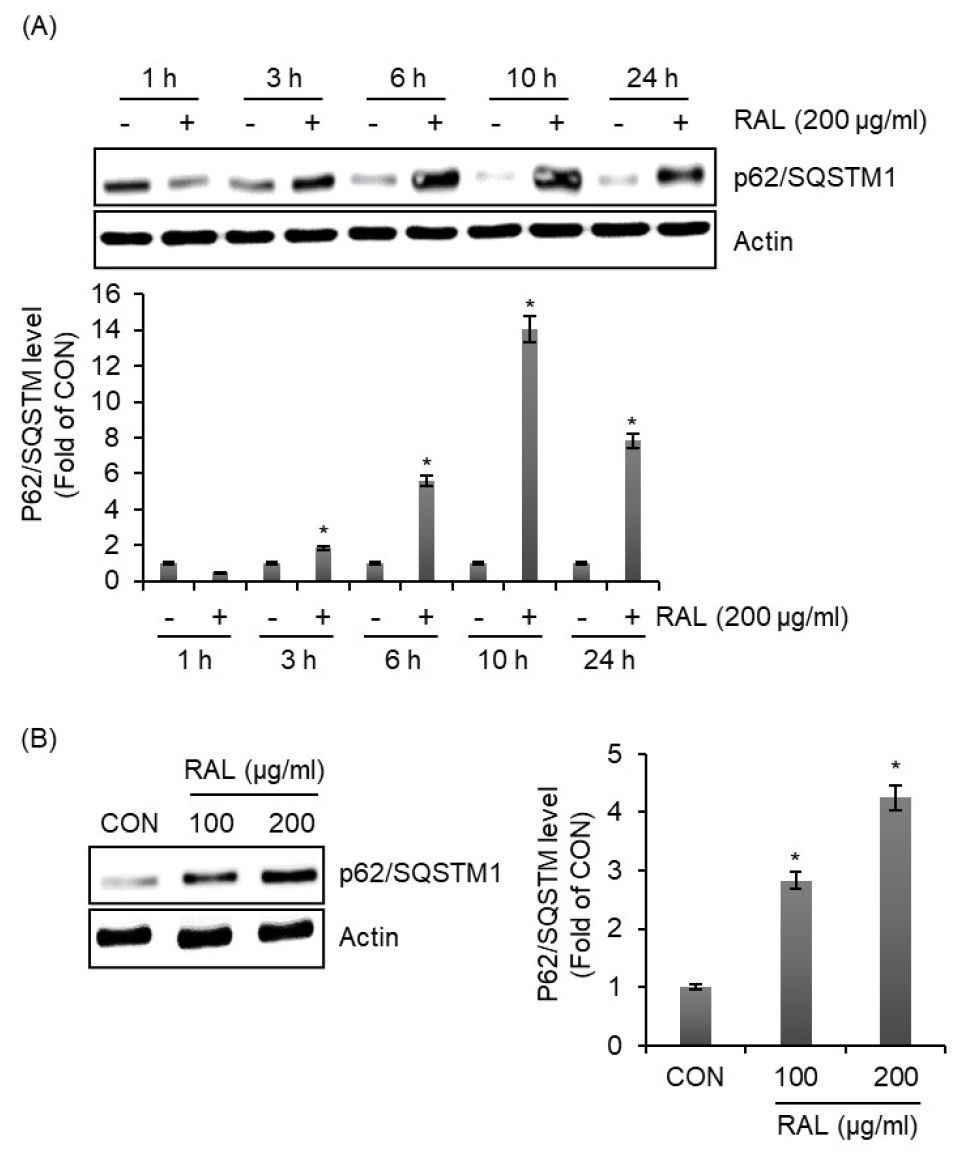
Fig. 2.
The effect of RAL on the expression of p62/SQSTM1 in RAW264.7 cells. (A) RAW264.7 cells were treated with RAL (200 ㎍/mL) for the indicated times. (B) RAW264.7 cells were treated with RAL (200 ㎍/mL) at the indicated concentrations for 6 h. The level of p62/SQSTM1 was analyzed by Western blot analysis.
Toll-like receptor 4 (TLR4)의 인가목 잎 추출물에 의한 대식세포의 자가포식 유도에 미치는 영향
대식세포는 Toll-like receptors (TLRs)을 통해 외래 병원체를 인식하면서 인체의 면역반응을 개시한다고 알려져 있고 이러한 과정은 인체의 면역적 방어에 매우 중요하다고 보고되고 있다(Chou et al., 2020). 최근 보고에 따르면, TLRs는 자가포식을 유도하여 대식세포의 항원 처리 및 항원 제시 기능을 향상시키는 것으로 보고되었다(Chou et al., 2020). 게다가 TLRs는 자가포식 과정에서 중요한 자가포식소체의 형성에도 관여한다고 알려져 있다(Wu and Lu, 2020).
인가목 잎 추출물 매개 p62/SQSTM1 단백질 수준 증가에 TLR2와 TLR4가 관여하지 조사하기 위해 TLR2와 TLR4 억제제 존재 하 인가목 잎 추출물을 처리하고 Western blot 분석으로 p62/SQSTM1 단백질 발현 수준을 확인하였다. 그 결과, C29에 의한 TLR2의 억제는 인가목 잎 추출물 매개 p62/SQSTM1 단백질 수준 증가에 아무런 영향이 없었으나, TAK-242에 의한 TLR4의 억제는 인가목 잎 추출물 매개 p62/SQSTM1 단백질 수준 증가를 주목할 만하게 감소시켰다(Fig. 3). 이러한 결과는 인가목 잎 추출물 매개 p62/SQSTM1의 증가는 TLR4에 의존적이라고 판단된다.

Fig. 3.
The effect of TLR2/4 on the RAL-mediated expression of p62/SQSTM1 in RAW264.7 cells. RAW264.7 cells were treated with RAL (200 ㎍/mL) in the absence or presence of C29 (TLR2 inhibitor, 100 nM) or TAK-242 (TLR4 inhibitor, 5 ㎍/mL) for 6 h. The level of p62/SQSTM1 was analyzed by Western blot analysis.
TLRs 중 TLR4 의존적 자가포식은 자가포식소체를 형성하여 외래 병원체를 제거한다고 알려져 있다(Xu et al., 2007). 또한 TLR4 매개 자기포식 유도는 백신 보조제 개발을 위한 효과적인 표적으로 사용되고 있으며 자가포식 기능을 활성화할 수 있는 TLR4 작용제들은 임상에서 백신 보조제로 사용되고 있다(Steinhagen et al., 2011). 따라서, 인가목 잎 추출물의 TLR4 매개 대식세포 자가포식 유도는 인가목 잎 추출물이 백신 보조제 개발을 위한 잠재적 소재로 활용될 수 있음을 보여준다.
PI3K/AKT 신호전달의 인가목 잎 추출물에 의한 대식세포의 자가포식 유도에 미치는 영향
Mitogen-activated protein kinases (MAPK), nuclear factor kappa B (NF-κB) 또는 phosphoinositide 3-kinase/protein kinase B (PI3K/AKT)의 활성화는 TLR4 의존적 자가포식 유도에 필수적이라고 보고되고 있다(Puissant et al., 2012; Van der Vaart et al., 2014; Yazid and Hung-Chih, 2021). 그리하여 RAW264.7 세포에서 인가목 잎 추출물에 의한 TLR4 매개 자가포식 유도에 어떤 신호전달이 관여하는지 조사하였다. 그 결과, PD98059에 의한 extracellular signal-regulated kinase 1/2 (ERK1/2) 억제, SB203580에 의한 p38의 억제 그리고 11-7082에 의한 NF-κB의 억제는 인가목 잎 추출물에 의한 p62/SQSTM1 단백질 수준 증가에 아무런 영향이 없었다(Fig. 4A). 그러나 SP600125에 의한 c-Jun N-terminal Kinase (JNK) 억제와 LY294002에 의한 PI3K/AKT 억제는 인가목 잎 추출물 매개 p62/SQSTM1 단백질 수준 증가를 주목할 만하게 감소시켰다(Fig. 4A). 이러한 결과는 인가목 잎 추출물 매개 p62/SQSTM1 단백질 수준 증가는 JNK 및 PI3K/AKT 신호전달의 활성화에 기인한 것으로 판단된다. 그리하여 인가목 잎 추출물이 JNK와 PI3K/AKT 신호전달을 활성화하는지 조사하였다. 그 결과, 인가목 잎 추출물은 JNK, PI3K, 그리고 AKT의 인산화를 유도하였고(Fig. 4B), 이것은 인가목 잎 추출물이 JNK와 PI3K/AKT 신호전달의 활성화를 유도한다는 것을 보여준다. 그리고 TAK-242에 의한 TLR4 억제 시, 인가목 잎 추출물 매개 JNK, PI3K 및 AKT의 인산화가 감소하였다(Fig. 4C). 본 결과를 미루어 볼 때, 인가목 잎 추출물에 의한 p62/SQSTM1 단백질 수준 증가는 TLR4 의존적 JNK와 PI3K/ATKT 신호전달 활성화에 기인한 것으로 판단된다.

Fig. 4.
The effect of MAPK, NF-κB, and PI3K/AKT signaling on the RAL-mediated expression of p62/SQSTM1 in RAW264.7 cells. (A) RAW264.7 cells were pretreated with PD98059 (ERK1/2 inhibitor, 40 μM), SB203580 (p38 inhibitor, 40 μM), SP600125 (JNK inhibitor, 40 μM), BAY 11-7082 (NF-κB inhibitor, 20 μM), or LY294002 (PI3K inhibitor, 40 μM) for 2 h and then co-treated with RAL (200 ㎍/mL) for 6 h. (B) RAW264.7 cells were treated with RAL (200 ㎍/mL) for the indicated times. (C) RAW264.7 cells were treated with TAK-242 (TLR4 inhibitor, 5 ㎍/mL) for 2 h and then co-treated with RAL (200 ㎍/mL) for 30 min. The protein level was analyzed by Western blot analysis.
적 요
본 연구에서 인가목 잎 추출물이 RAW264.7 세포에서 면역자극인자의 생성을 증가시키고 포식작용의 활성화를 유도한다는 것을 확인하였다. 게다가 인가목 잎 추출물은 TLR4 의존적 JNK와 PI3K/AKT 신호전달 활성화를 통해 자가포식을 유도한다는 것을 확인하였다. 대식세포의 활성화와 자가포식은 선천면역반응과 후천면역반응을 향상시킬 수 있는 주요 전략 중 하나이기 때문에 인가목 잎 추출물은 인체의 면역반응을 증가시킬 수 있는 건강기능식품이나 보조제로 활용될 수 있을 것으로 생각된다. 그러나 본 연구에서는 작용기전을 구명하기 위해 관련된 단일억제제를 사용하였기 때문에 명확한 작용기전 구명을 위해 다양한 억제제 또는 siRNA를 활용한 추가적 연구가 필요하다고 판단된다.
