

서 언
재료 및 방법
조사 대상지 및 현장조사
분석 방법
결 과
종면적곡선 및 적정 군집수 선정
식생 군집 분류
군집 간 환경인자 특성 분석
종조성 분석
Ordination 및 종간 상관관계
지표종 분석
고 찰
적 요
서 언
21세기 동안 인류는 자연생태계와 더불어 상호영향을 받고 생존해 왔으며, 이는 육지의 75%와 대부분의 해양과 관련이 있다(Lewis et al., 2017). 최근까지 인류가 시행해온 생태계 보전, 즉 현지내 보전의 핵심활동은 바로 보호구역의 법적 지정이다(CBD, 2024).
보호구역이란 “명확하게 정의된 구역으로 장기적인 자연 생태계 보전을 달성하기 위해 국가 및 국제적으로 인정되어 관리되는 지역”으로 정의한다. 보호구역은 지구 전체를 기준으로 육지 및 내륙의 14.7%에 포함된다고 보고되고 있으며, 최근 이것의 확대 지정은 기후위기 및 펜데믹에 따른 인류의 생존권을 보장하는 국가적 정책옵션으로 자리잡고 있다(Bingham et al., 2019).
보호구역은 IPBES (The Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services)에서 수행된 생태계 생물다양성 평가와도 관련이 있다(IPBES, 2016). 보호구역의 유지 및 확대지정은 생물다양성에 관한 전략적 계획을 통해 196개 국가가 합의했던 20개 목표 중 하나인 Aichi 생물다양성 목표 11의 핵심이다. 이는 2020년까지 육지와 내륙 수면의 17% 및 해양의 10%를 보호구역으로 확대하는 것을 목표로 하며, 효과적 관리 등 중요한 생물다양성 보전의 질적 향상을 충족시키는 것이다(CBD, 2024).
생물다양성 보전의 경향성은 최근 제15차 생물다양성협약의 당사국총회(CBD COP-15)에서 ‘쿤밍-몬트리올 글로벌 생물다양성 프레임워크(Kunming-montreal Global Biodiversity Framework) 채택에서 이를 뒷받침하고 있다. 그 중 Action target 3이 핵심목표로 ‘2030년까지 전 지구적 육상생태계 및 해양생태계 최소 30%를 보호지역으로 효과적으로 보전과 관리한다’이다(CBD, 2024; Heo and Park, 2023). 이러한 생태계 보호구역의 진행에 따라 전세계적 규모의 보호구역 관리 및 분석은 WDPA (the World Database on Protected Areas)를 기반으로 수행하고 있다(WDPA, 2024).
WDPA는 육지 및 해양 보호구역에 관하여 국제적으로 관할하는 데이터베이스이다. WDPA는 유엔 환경 계획과 국제자연보전연맹(IUCN, the International Union for Conservation of Nature) 사이의 공동 프로젝트이며, 유엔 환경 계획 세계 보전 모니터링 센터(UNEP-WCMC, UN Environment Programme World Conservation Monitoring Centre)가 정부, 비정부 기관, 학계 및 산업과의 협력을 통해 운영되고 있다. 이는 보호구역 데이터를 공유함으로써 다양한 보전 목표에 대한 분석과 평가와 관련한 글로벌 생물다양성 협약을 이행하는 것이다(WDPA, 2024).
이러한 국제적 움직임에 발맞추어 산림청 등 정부기관에서는 ‘제3차 산림유전자원보호구역 지역관리계획(2023~2027)’을 발표하였다. 이는 기후변화 등 외부적 위협요인으로부터 산림유전자원을 보호하고 산림생물다양성을 유지·증진하기 위한 보호구역 확대와 효율적인 관리, 지속가능한 이용체계 마련 등을 수행하고 있다(KFS, 2023). 이러한 국내‧외적 움직임은 보호구역의 확대 지정과 더불어 현재 지정된 보호구역 유지 및 관리의 중요성도 더불어 시사하는 것이다. 이를 위하여 보호구역의 식생자료를 구축하고 식생과 환경인자간의 상호영향을 파악하는 것은 산림보전 및 관리에 있어 기본적인 연구이다(Newton, 2007; Wilson, 1992).
식생이란 특정 서식처 내 식물종의 모임이고 이들이 제공하는 피복 정도를 의미한다. 특히 산림식생의 경우 기후, 토양, 지형조건 등과 같은 다양한 환경 요소에 영향을 미치는 생태계의 중요한 구성 요소이다. 식생 연구는 식물종과 군집 분포, 풍부성과 동태를 분석하는 것이며, 이는 식물 군집이 환경요인과 상호작용하고 변화에 따라 어떻게 반응하는지 이해하는데 필수적이다(Barbour et al., 2004). 이를 기반으로 식생 연구는 보호구역의 생물다양성 유지 및 관리, 그리고 보전을 위한 기본적인 연구를 수행한다. 그 이유는, 식생 연구는 다양한 생물종의 서식지에 해당하므로 이들의 동태와 분포를 파악하기 위한 상관성을 점검하는 것이며, 이를 관리하기 위한 과학적 기반을 제공하기도 한다. 즉, 식생연구는 생태계 보전의 장기적 지속 가능성을 보장하기 때문이다(Barbour et al., 2004; Newton, 2007).
한편 방태산(1,445 m)는 국내 산림유전자원보호구역에 포함될 뿐만 아니라 WDPA에 등재된 곳으로, 국제적인 생태계 보호구역으로서의 가치가 있는 지역이다. 방태산은 강원도 인제와 홍천군에 경계를 이루고 있으며, 방태산은 우리나라 생물다양성의 보고인 설악산과 오대산의 중앙에 위치하여 종분포 이입과 유출의 이동통로의 의미가 있어 식물지리학적 가치가 뛰어난 지역이다. 또한 방태산은 아고산 식생대에 분포하고 있어 국내에 협소한 서식처이며, 북방계 식물의 피난처로서 역할을 한다. 그러나 최근 기후위기로 인하여 아고산 식생대는 급속한 지구환경변화로 인한 쇠퇴가 우선적으로 일어나는 지역으로 면밀한 식생 구조연구가 필요하다(Kong, 2002; Park et al., 2019; Park et al., 2020). 현재 방태산의 식물생태계와 관련된 선행연구로는 식물상에 대한 기초자료는 구축되어 있으며(KNA, 2013; Lee et al., 1997), Kil et al. (1995)에 의하여 방태산 북사면의 식생군집구조가 연구되었다. 그러나 특정 사면위주의 식생 군집을 파악하였으며, 군집분류, 군집간 유사성과 종다양성에 대한 논의가 이루어졌으나 식생군집간 환경인자와 영향을 미치는 요인 등 상관관계에 대한 연구는 미흡한 실정으로 이에 대한 보완이 필요하다.
본 연구의 목적은 WDPA에 속하며, 산림유전자원보호구역에 속하는 방태산 일대의 식생군집과 생태계요인과의 상호관계를 확인하여 보호구역의 다각적 보전을 위한 기초자료를 제공하는 것이다.
재료 및 방법
조사 대상지 및 현장조사
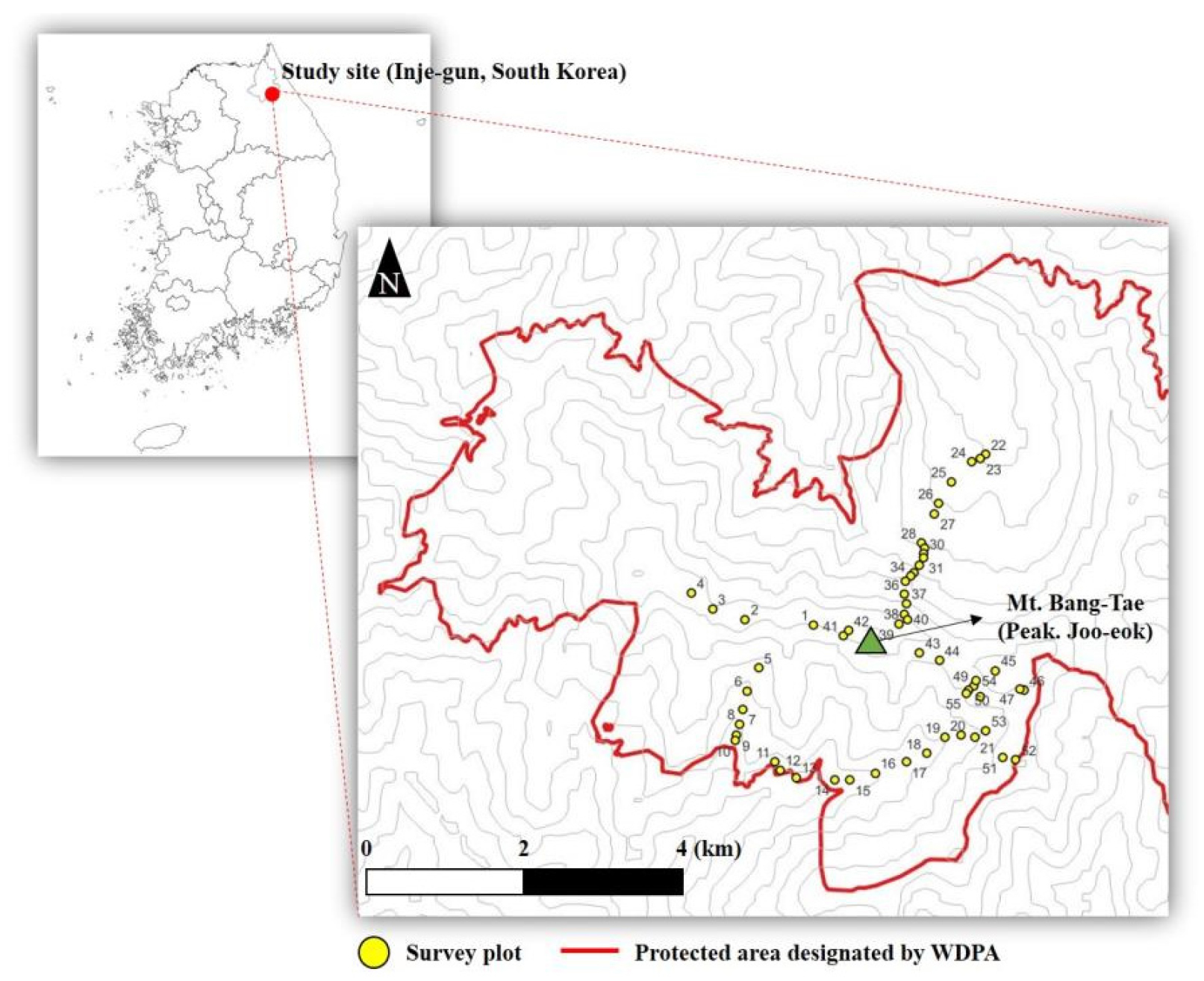
본 연구대상지인 방태산은 WDPA에 속하면서 국내 산림유전자원보호구역에 포함되는 곳으로, 주변의 산지로는 구룡덕봉(1,388 m), 가칠봉(1,156 m), 응복산(1,241 m) 등이 포함되어 산새가 험하며 암반지대가 많으며, 아고산 식생대에 속하는 산지이다(KNA, 2013). 방태산을 중심으로 장거리의 능선과 깊은 골짜기가 자리잡고 있으며, 내륙성 산지에 속한다. 또한 적가리골, 골안골 등 수려한 자연경관을 보유한 지역이며, 지형이 다양하여 생물다양성의 보호 가치가 높은 지역이다(Kil et al., 1995).
행정구역상 위치는 강원특별자치도 인제군에 속해 있으며, 본 식생 조사지는 위도 37°52′16.91″~37°54′29.91″, 경도 128°21′48.00″~127°24′39.37″의 공간적 범위에 속해있다(Fig. 1).
기상청(2024)에서 제공한 인제군의 30년간(1994~2023) 연평년값을 기준으로 한 기상관측소 데이터를 분석한 결과 연평년기온은 10.5℃, 연평년최저기온 5.2℃, 연평년최고기온 16.8℃, 연평년강수량 1,227.9 ㎜, 평균풍속 1.8 m/s, 평균상대습도 68.0%으로 기록되었다(KMA, 2024). 전형적인 산악지형의 기후 특징으로 나타나고 있으며, 해발고도가 상대적으로 높아 일교차가 심하며 하계에는 서늘하고, 동계에는 적설량이 높아 추운 지역이다.
WDPA지정된 보호구역을 중심으로 총 55개 식생조사구를 설치하였다. 식생조사구 내 층위구조를 확인하여 20 m×20 m의 정방형 식생 조사구로 하나의 식생 조사구는 400㎡로 설정하였다. 식생 조사 방법은 Braun-Blanquet (1964)에 따른 교목층, 아교목층, 관목층과 초본층을 구분하여 식물을 동정한 뒤, 피도와 밀도를 조사하였다. 조사기간은 2019년 3월에서 2022년 10월까지 수행되었다.
식생조사구 내 출현한 식물의 동정은 Korean Fern Society (2005), KNA (2020)를 참조하였다. 학명 및 국명은 국가표준식물목록(KNA, 2020)에 준하였다.
분석 방법
식생군집 분류의 전단계 분석으로 해당 식생 조사구의 설치가 적정한지에 대한 검증을 위하여 종면적곡선(Species area curves) 분석을 수행하였으며, 이는 Jacknife 방법에 의한 종수 추정에 의한 종면적곡선은 비모수적 표본수가 약 100표본 내외로 소수일 경우 사용되며, 실측 데이터에 대한 결과를 통하여 신속하고 간결하게 적정 조사구 수 평가가 가능하다(Chao et al., 2005; Efron and Stein, 1981). 적정 클러스터 개수를 파악하기 위하여 각 군집에서 통계적 유의성이 인정되는(P<0.05) 가장 많은 지표종과 그 들의 통계값에 의한 P-value의 평균이 가장 낮은 값으로 선정된 군집수를 적정 군집 개수로 정의한다(McCune and Grace, 2002; McCune and Mefford, 2006).
식생군집 분류는 클러스터 분석을 이용하여 dendrogram을 작성하였으며, 거리척도는 유클리디안 거리척도를 이용하였다. 식생군집 분류에 필요한 종조성 데이터는 식생조사에서 수행된 피도와 밀도계급의 중앙값을 이용하였다.
군집 간 환경조건의 차이를 확인하기 위하여 환경요인에 따른 군집별 일원배치분산분석(one-way ANOVA)을 사용하였으며, 사후검증은 Duncan‘s method를 이용하였다.
군집별 식물종의 우점정도는 종조성 데이터(상대피도, 상대밀도)의 중요치를 분석하였으며, Curtis and McIntosh (1951) Shannon’s index를 통하여 종다양성 평가를 시행하였다(Shannon and Weaver, 1949).
종조성의 2차원상 배열과 환경인자간 상관관계를 분석하기 위하여 NMS (Non-metric Multidimensional Scaling) ordination을 수행하였다 (cut off R2=0.3). 환경인자는 해발고도, Northness, 경사, 노암율, TPI (Topographic Position Index), TWI (Topographic Wetness Index), 종풍부도, 균재도, 종다양도 그리고 중요치 분석 후 상위 10% 이내에 속하는 식물종(Pinus sp., Acer sp., Quercus sp., Sasa sp., Abies sp., Euonymus sp., Magnolia sp., Drypteris sp., Carpinus sp., Calamagrostis sp., Tilia sp., Betula sp., Lindera sp., Tripterygium sp., Rhododendoron sp., Deutzia sp.)의 속(genus)의 피도값으로 선정하여 총 25개를 분석에 이용하였다. Northness는 현장에서 취득한 사면방위를 라디안 각도 변환 후, 코사인 삼각함수를 적용하여 –1에서 1사이 수치로 환산하였다. 해당 식생 조사구의 Northness 수치가 1에 가까울수록 북사면에 해당하며 –1에 근접할 수록 남사면에 위치함을 의미한다(Park et al., 2023).
NMS ordination 상 식물종의 상관관계를 파악하기 위하여 Strong association을 파악하여(cut off R2=0.3) 2차원상에 도출하였다(McCune and Grace, 2002; McCune and Mefford, 2006).
TWI (Topographic Wetness Index)와 TPI (Topographic Position Index)는 DEM (Digital Elevation Model)을 기반으로 추출하였다. 획득한 DEM 원자료는 국토지리정보원(2024)에서 제공하는 자료를 이용하였으며, 이를 기반으로 TWI 및 TPI를 추출하였다. TWI는 특정 지역의 토양 수분조건의 척도가 되며(Petroseli et al., 2013), TPI는 음수일수록 계곡부(오목한 지형)이며, 양수일수록 능선부(볼록한 지형)로 해석되어 지형의 특성을 파악하는 간접적인 지수가 된다(Newman et al., 2018).
군집별 종조성의 구분에 영향을 미치는 식물종을 파악하기 위하여 지표종 분석(Indicator Species Analysis)을 수행하였다. 이를 위하여 각 군집별 MRPP-test (Multi-Response Permutation Procedures)를 통하여 군집 별 종조성의 이질성 검증하였다. MRPP-test는 군집 분류 해석을 위한 식생 데이터에서 이질성 검증을 하는 적합한 분석방법이다(McCune and Grace, 2002; McCune and Mefford, 2006). MRPP-test는 T통계량과 A척도를 나타내는데, T통계량이 낮을수록 군집 내 유사함을 의미하며, A척도가 높을수록 비교 군집 간 이질성이 높음을 의미한다(McCune and Mefford, 2006). 지표종은 각 군집별로 통계적 유의차가 0.05 미만인 종으로 선정하였으며, 지표값(Indicator value)은 4,999회의 몬테 카를로 시뮬레이션(Monte Carlo Simulation)을 이용하여 계산하였다(Dufrêne and Legendre, 1997).
식생 데이터 분석프로그램은 SPSS Statistics (ver. 21.0), PC-ORD (ver. 7.0)를 이용하였으며, GIS 분석은 ArcGIS (ver. 10.8)를 이용하였다.
결 과
종면적곡선 및 적정 군집수 선정
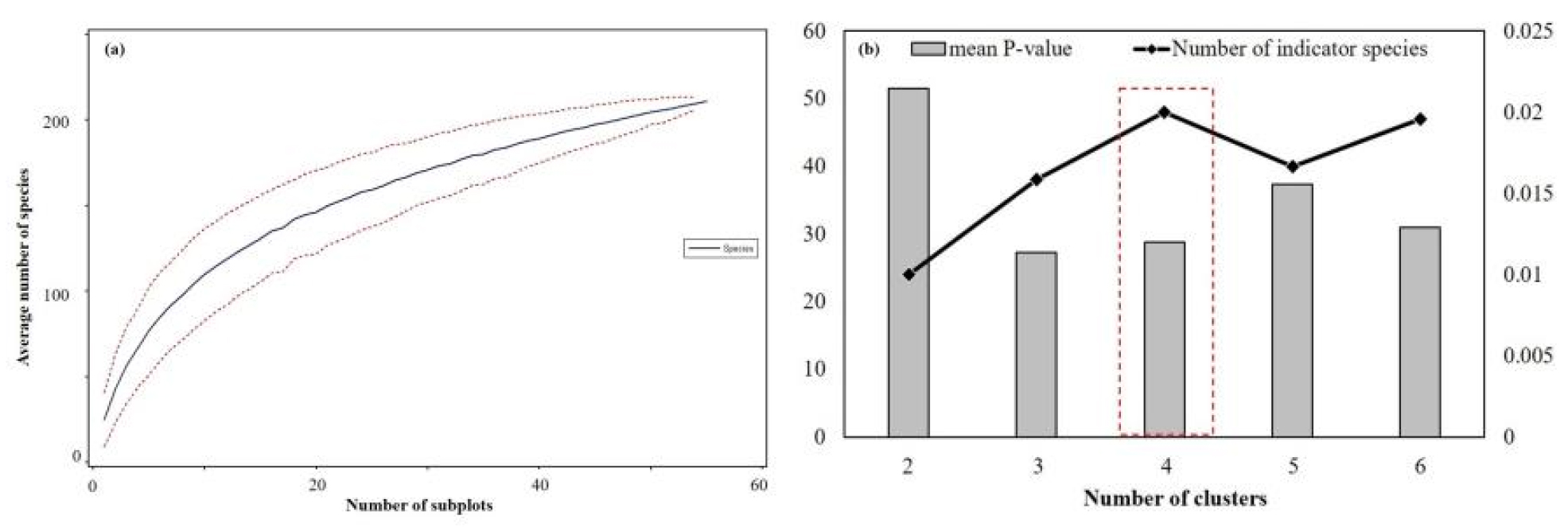
본 연구 분석의 적절성 및 식생 군집의 적정 수를 파악하기 위한 분석결과는 Fig. 2와 같다. 본 식생 조사구 내 출현한 식물종수는 60과 137속 184 종 2아종 21변종 4품종으로 총 211분류군으로 조사되었다.
본 연구 분석의 조사구수가 생태계 분석에 적합한지에 대한 검증을 위하여 종수추정에 따른 종면적 곡선(Species area curve)을 작성하였다(Fig. 2-a). 분석 결과 55개소의 표준지에 대하여 조사구 증가에 따른 종풍부도의 기울기가 0에 수렴하는 것으로 나타나 생태 데이터 분석에 적정한 조사구 수임을 증명하였다(McCune and Mefford, 2006).
또한 적정 군집수를 선정하기 위하여 군집 수를 2개~6개로 비교하여 적정 클러스터 군집수를 검증하였다(Fig. 2-b). 분석 결과 방태산의 적정 식생 군집수를 확인하기 위하여, 식생 군집 분류에 따른 지표종의 수와 P-value의 평균값을 계산하였다. 지표종 선정은 Indicator value에 따른 통계값이 P<0.05 이하인 식물로 정의한다(McCune and Grace, 2002). 분석결과 식생 군집이 4개일 때 적정 군집분류 수로 분석되었다. P-value의 값은 3개 군집에서 가장 낮았으나(P=0.0113), 차순위 군집 수는 4개로 분석되었고(P=0.0120), 지표종수는 4개 군집에서 가장 많은 것으로 분석되었다(48 taxa). 종합적으로 4개의 군집수에서 낮은 P-value값과 지표종수로 판단되어 본 연구분석의 적절한 식생 군집수로 선정하였다.
식생 군집 분류
Cluster dendrogram을 통한 식생 군집 분류를 수행한 결과 Fig. 3과 같다. 총 4개 군집으로 분류되었으며, 각 군집 간 종조성의 이질성 판단을 통한 군집 분류의 적합성을 MRPP-test를 통하여 검증하였다(Table 1). 검증결과, 각각의 군집 내 통계적 유의성이 인정되어 적절한 군집으로 분류된 것으로 검증되었다(P<0.001). 4개 군집의 명명은 종조성에 따른 중요치에 따라 선정하였으며, 그 결과 당단풍나무-고로쇠나무 군집(C1), 신갈나무 군집(C2), 분비나무 군집(C3) 그리고 조릿대-당단풍나무 군집(C4)로 분류되었다. C1은 대부분 계곡부 또는 사면하 부분에서 주로 분포하고 있었으며, C2와 C4는 대부분 사면 중‧상부에 위치하고 있었다. C3는 멸종위기 고산침엽수인 분비나무림으로 사면상부 또는 능선부에 위치하고 있었다.
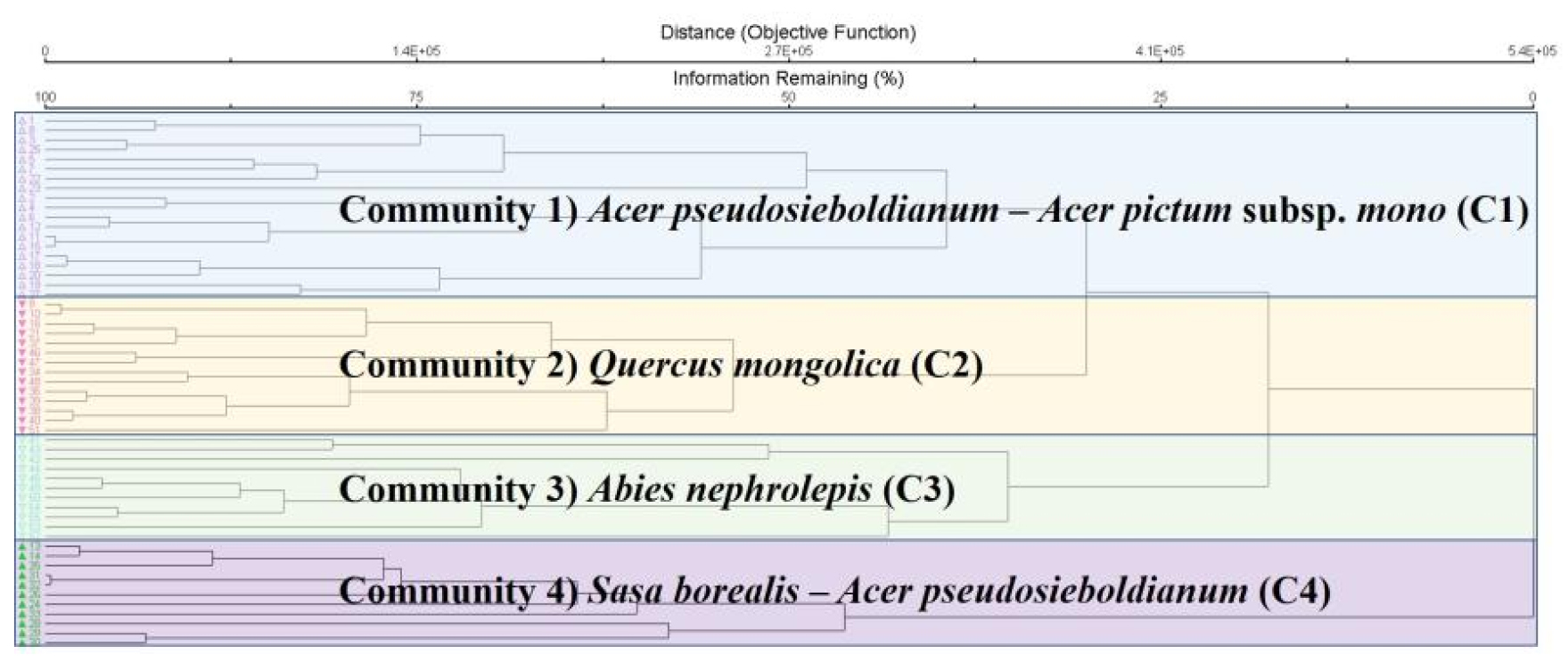
Table 1.
Results of MRPP-test for each community in Mt Bangtae.
| Compared communities | Tz | Ay | p-value |
| C1 vs C2 | -11.7048 | 0.269278 | <0.001 |
| C1 vs C4 | -13.3832 | 0.342378 | |
| C1 vs C3 | -16.2313 | 0.425752 | |
| C2 vs C4 | -10.8768 | 0.321229 | |
| C2 vs C3 | -9.65158 | 0.270408 | |
| C4 vs C3 | -12.7779 | 0.476608 |
군집 간 환경인자 특성 분석
각 군집 간 환경인자의 특성을 확인하기 위하여 각 인자별 one-way-ANOVA를 이용하여 확인한 결과 Table 2와 같다. 해발고도의 경우 C3에서 1,249.1±21.8 m로 가장 높게 나타났으며, Northness는 C4에서 0.2844±0.2418로 타 군집에 비하여 북사면 경향이 있는 것으로 나타났으며, 그 수치가 크지는 않은 것으로 나타났다. 국내 Abies 속에 속하는 분비나무의 경우 국내 대부분 서사면을 제외하고 북쪽사면에 주로 분포하고 있으나(Park et al., 2022), 본 분석에서는 음의 값으로 나타나 남사면에 가까운 경향으로 나타났다. 이는 강원 남부지역 분비나무림의 순림형태의 경우 남사면에 주로 분포하는 선행연구와 유사한 연구결과로 나타났다(Shin and Oh, 2022).
경사도의 경우 C3에서 33.4±2.3°로 가장 가파른 경사도로 나타났으며, 암석노출도도 69.5±3.1%로 나타나 타 군집에 비하여 험준하며 척박한 지형으로 나타났다.
지형수분지수(TWI)의 경우 C1에서 8.887±0.623으로 나타나 가장 높게 나타나 지형특성에 따른 수분함량이 가장 양호한 지역으로 분석되었다. 지형지수(TPI)의 경우 C4에서 5.244±3.902로 가장 높게 나타났다.
각 군집별 환경인자는 해발고도 및 암석노출도에서는 P-value가 0.001미만으로 나타났으며, Northness, 경사도, TWI의 경우에는 P-value가 0.05이하로 나타나, 각 군집별 환경 인자의 차이가 있음을 의미한다. 지형지수는 표준오차가 높고 편이며, 각 조사구마다 균일한 지수를 나타나진 않아 통계적 유의성이 인정되지 않은 것으로 나타났다(P >0.05).
Table 2.
Characteristic of environmental factors for each community using one-way ANOVA.
종조성 분석
방태산 일대 중요치를 분석한 결과는 Table 3과 같다. C1에서는 당단풍나무(7.02)→고로쇠나무(4.88)→복장나무(4.74)→생강나무(4.35)→물참대(3.39) 등의 순으로 나타났으며, C2는 신갈나무(11.08)→당단풍나무(7.00)→나래회나무(4.45)→분비나무(4.38)→고로쇠나무(4.38) 등의 순으로 조사되었다. C3는 분비나무가 15.06으로 타 수종보다 월등히 높은 중요치로 나타났으며, 그 뒤로 실새풀(6.11)→부게꽃나무(5.93)→고로쇠나무(4.50)→관중(4.06) 등의 순으로 나타났다. C4는 조릿대와 당단풍나무가 각각 17.89, 15.45로 높은 중요치로 나타났으며, 그 뒤로 신갈나무(12.47)→전나무(3.73)→피나무(3.53)→까치박달(3.29)→잣나무(2.99) 등의 순으로 분석되었다.
Table 3.
Importance percentage for 4 vegetation communities classified in Mt. Bangtae.
각 군집별 종다양성 분석 결과 Table 4와 같다. C4에서 종풍부도(SR), 균등도(Even.) 및 종다양도(H’)가 모두 낮게 나타났으며, 각 군집별 분석항목의 통계적 유의차가 인정되는 것으로 나타났다(P<0.05).
Table 4.
Results of species diversity for 4 vegetation communities.
| Contents | C1 | C2 | C3 | C4 | P |
| SRz | 25.6±1.3b | 27.6±2.7b | 27.2±2.1b | 18.5±1.8a | 0.016 |
| Even.y | 0.902±0.005b | 0.893±0.008b | 0.887±0.010b | 0.858±0.010a | 0.005 |
| H'x | 2.907±0.044b | 2.916±0.084b | 2.909±0.101b | 2.471±0.094a | <0.001 |
Ordination 및 종간 상관관계
식생 군집을 NMS ordination을 통하여 종조성의 이질성 및 환경간의 상관관계를 파악하기 위하여 2차원상 배열 분석한 결과는 Fig. 4와 같다. 1축과 2축의 설명력은 각각 0.409, 0.267로 종합설명력 0.677로 분석되었다. 각 식생군집의 교집합 점이 최소로 남아 있어 각 군집별 종조성의 이질성이 육안으로 확인되었다. 특히, C3는 타 군집에 비하여 종조성이 이질적인 것으로 나타났다.
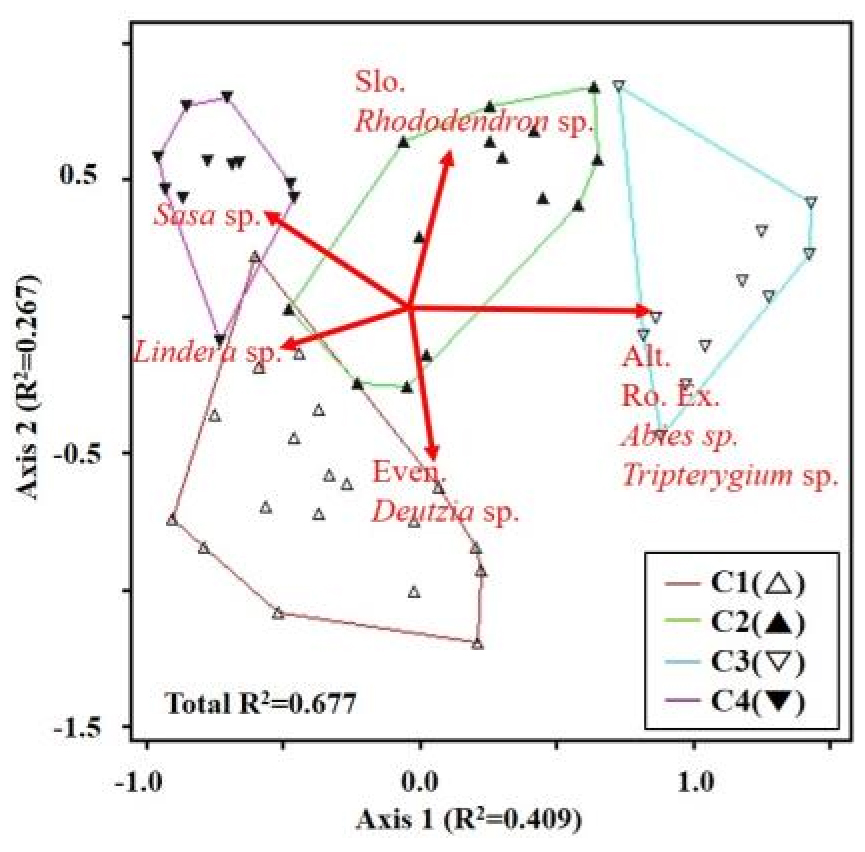
식생 군집과 환경인자와의 상관관계를 살펴보면, C1의 경우 균등도와 Deutzia sp. 그리고 Lindera sp. 피도와의 상관관계를 나타내고 있었으며, C2의 경우 경사도와 Rhododendron sp.와의 피도와 상관관계로 나타났다. C3는 해발고도, 암석노출도, Abies sp. 및 Tripterygium sp.의 피도와 관계가 있는 것으로 나타났으며, C4는 Sasa sp.와의 상관관계로 나타났다.
NMS ordiantion을 통한 종별 strong association 분석을 통한 종간 상관관계를 파악하여 영향성을 확인한 결과는 Fig. 5와 같다. 분석 결과, 오미자(x1)-눈빛승마(x2), 나래회나무(x3)-큰개별꽃(x4), 음나무(x5)-조릿대(x6), 딱총나무(x7)-물참대(x8)-십자고사리(x9)로 나타났다. 그리고 종간 상관관계가 광범위한 미역줄나무(x10)-분비나무(x11)-시닥나무(x12)-사스래나무(x13)-퍼진고사리(x14)로 분석되었다. 분비나무의 경우 C3의 대표 군락으로, 미역줄나무와 강한 영향 관계로 나타나는 것으로 나타났으며 그 밖에 내음성 수종인 시닥나무와, 사스래나무와도 동일한 생태적 지위 내에서 상호 영향을 미치는 것으로 나타났다. C3군집은 분비나무 군집으로, 해당 군집의 종조성이 높은 해발고도와, 암석노출도 그리고 미역줄나무와의 상관관계가 나타났다. 해당 군집이 방태산 산림유전자원보호구역의 아고산 식생대를 대표하는 군집이다. 해당 군집은 서식환경이 척박하며, 풍해에 의한 영향이 많아 아고산 침엽수 및 맹아갱신한 활엽수의 1차적 물리적 피해가 대부분이며, 이후 발생한 숲틈을 통하여 2차적 생리적 피해가 발생하는 지역이다(Hasegawa and Mori, 2007; Han et al., 2012; Kim et al., 2019; Park et al., 2021).
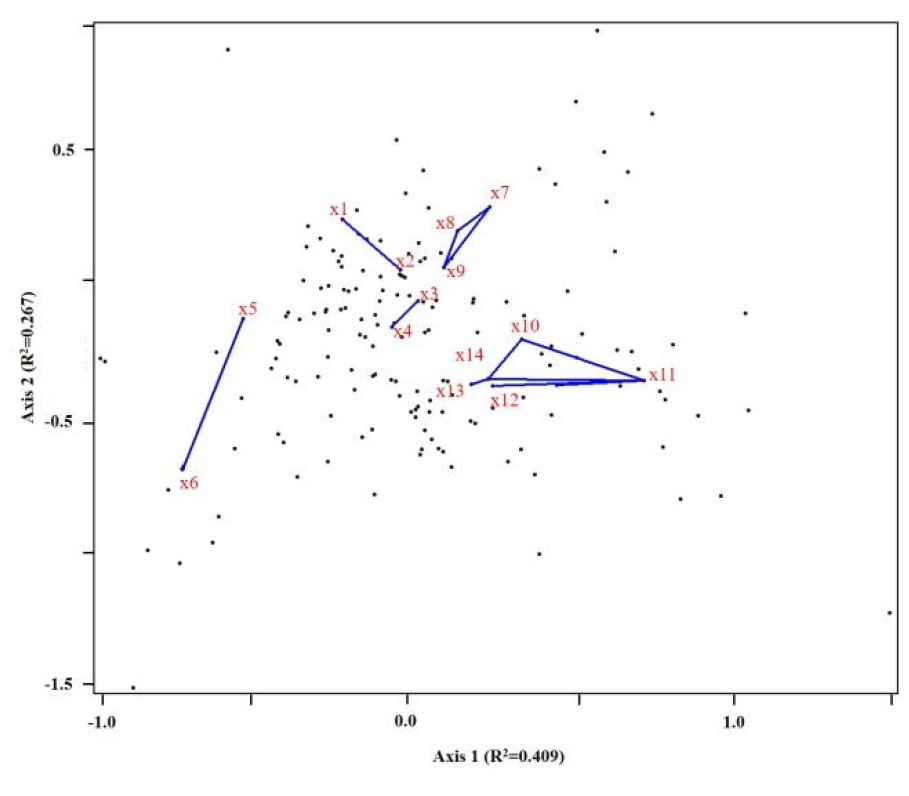
Fig. 5.
Distribution of species (black dots) and interspecies correlation(blue line, strong association cut off R2=0.3) on 2-dimension space (x1: Schisandra chinensis, x2: Cimicifuga dahurica, x3: Euonymus macropterus, x4: Pseudostellaria palibiniana, x5: Kalopanax septemlobus, x6: Sasa borealis, x7: Sambucus williamsii var. coreana, x8: Deutzia glabrata, x9: Polystichum tripteron, x10: Tripterygium regelii, x11: Abies nephrolepis, x12: Acer komarovii, x13: Betula ermanii, x14: Dryopteris expansa).
지표종 분석
지표종 분석 결과 Table 5와 같다. C1의 경우 오미자(51.0), 고추나무(40.9), 복장나무(39.9), 눈빛승마 등 12분류군으로 나타났다. C2는 단풍취(45.2), 지리대사초(30.8), 꼭두서니(26.3), 철쭉(26.0), 은분취(21.4)로 5분류군으로 분석되었다. C3의 경우 분비나무(80.7), 실새풀(75.0), 부게꽃나무(64.7), 주목(58.5), 마가목(58.1), 붉은병꽃나무(52.3) 등의 순으로 나타나 총 23분류군으로 가장 많은 지표종이 선정된 군집으로 분석되었다. C4는 조릿대(86.5), 쪽동백나무(62.5), 당단풍나무(53.3), 신갈나무(49.8) 등으로 전체 8분류군으로 지정되었다.
Table 5.
Results of indicator species analysis for each community.
| Contests | Scientific name / Korean name | IVz | Py |
| C1 | Schisandra chinensis / 오미자 | 51.0 | 0.0016 |
| Staphylea bumalda / 고추나무 | 40.9 | 0.0020 | |
| Acer mandshuricum / 복장나무 | 39.9 | 0.0058 | |
| Cimicifuga dahurica / 눈빛승마 | 38.2 | 0.0074 | |
| Morus bombycis / 산뽕나무 | 37.2 | 0.0036 | |
| Lindera obtusiloba / 생강나무 | 33.0 | 0.0218 | |
| Rubus crataegifolius / 산딸기 | 26.3 | 0.0102 | |
| Pinus densiflora / 소나무 | 26.3 | 0.0126 | |
| Corylus heterophylla / 개암나무 | 25.8 | 0.0222 | |
| Aster scaber / 참취 | 25.3 | 0.0212 | |
| Carex humilis var. nana / 가는잎그늘사초 | 23.6 | 0.0336 | |
| Ulmus davidiana var. japonica / 느릅나무 | 23.1 | 0.0438 | |
| C2 | Ainsliaea acerifolia / 단풍취 | 45.2 | 0.0010 |
| Carex okamotoi / 지리대사초 | 30.8 | 0.0074 | |
| Rubia akane / 꼭두서니 | 26.3 | 0.0152 | |
| Rhododendron schlippenbachii / 철쭉 | 26.0 | 0.0472 | |
| Saussurea gracilis / 은분취 | 21.4 | 0.0312 | |
| C3 | Abies nephrolepis / 분비나무 | 80.7 | 0.0002 |
| Calamagrostis arundinacea / 실새풀 | 75.0 | 0.0002 | |
| Acer ukurunduense / 부게꽃나무 | 64.7 | 0.0002 | |
| Taxus cuspidata / 주목 | 58.5 | 0.0002 | |
| Sorbus commixta / 마가목 | 58.1 | 0.0002 | |
| Weigela florida / 붉은병꽃나무 | 52.3 | 0.0006 | |
| Rhododendron mucronulatum / 진달래 | 50.4 | 0.0002 | |
| Dryopteris crassirhizoma / 관중 | 47.7 | 0.0002 | |
| Actinidia kolomikta / 쥐다래 | 45.5 | 0.0004 | |
| Arisaema amurense /둥근잎천남성 | 43.9 | 0.0008 | |
| Tripterygium regelii / 미역줄나무 | 42.9 | 0.0062 | |
| Syringa reticulata var. mandshurica / 개회나무 | 37.8 | 0.0026 | |
| Urtica angustifolia / 가는잎쐐기풀 | 36.4 | 0.0016 | |
| Dryopteris expansa / 퍼진고사리 | 35.4 | 0.0170 | |
| Syringa patula / 털개회나무 | 34.0 | 0.0034 | |
| Sambucus williamsii var. coreana / 딱총나무 | 32.7 | 0.0362 | |
| Pinus koraiensis / 잣나무 | 32.5 | 0.0218 | |
| Acer komarovii / 시닥나무 | 30.5 | 0.0216 | |
| Prunus padus / 귀룽나무 | 30.1 | 0.0320 | |
| Syringa wolfii / 꽃개회나무 | 29.8 | 0.0124 | |
| Pedicularis resupinata / 송이풀 | 28.0 | 0.0414 | |
| Agastache rugosa / 배초향 | 27.3 | 0.0132 | |
| Deutzia uniflora / 매화말발도리 | 23.3 | 0.0296 | |
| C4 | Sasa borealis / 조릿대 | 86.5 | 0.0002 |
| Styrax obassia / 쪽동백나무 | 62.5 | 0.0002 | |
| Acer pseudosieboldianum / 당단풍나무 | 53.3 | 0.0002 | |
| Quercus mongolica / 신갈나무 | 49.8 | 0.0006 | |
| Tilia amurensis / 피나무 | 33.5 | 0.0226 | |
| Sorbus alnifolia / 팥배나무 | 28.2 | 0.0182 | |
| Acer tegmentosum / 산겨릅나무 | 24.2 | 0.0216 |
고 찰
본 연구대상지인 방태산은 군집에 따른 지형환경 총 6개 환경인자 중 5개 항목에서 통계적 유의차가 인정되었다. 이러한 결과는 연구대상지는 지형 환경에 따른 식생 군집이 패치형 모자이크 구조를 이루고 있는 것으로 해석된다. 예를 들면, 다른 군집들과 비교하여 상대적 해발고도가 낮은 계곡부에는 C1 군집이, 높은 해발고도에 사면부에 위치하는 C2 군집, 해발고도 및 경사도가 크며, 능선부에 자리잡은 C3 군집, 그리고 해발고도가 낮은 사면하부는 C4 군집으로 이루어져 있다. 이러한 군집 경향은 황병산에서도 분비나무림의 군집구조가 환경 특성에 따른 종조성이 뚜렷하게 구분되는 모자이크식 패치를 나타내는 것과 유사한 결과를 도출하였다(Park et al., 2020). 본 연구 분석은 방태산 북사면을 중심으로 식생 군락을 분류한 결과에서도 사면부에서는 신갈나무 군락, 계곡부에는 Acer 속 및 들메나무 군락, 능선부에는 소나무과 군락이 우점하고 있다는 연구결과와 유사하였다(Kil et al., 1995). 본 연구결과에서도 C2(신갈나무군집)가 타 군집에 비하여 북사면에 위치하고 있는 것으로 분석되었다.
또한 선행연구에서 방태산일대는 신갈나무가 널리 우점한 지역으로 서술하고 있었으며(Kil et al., 1995) 신갈나무군락이 타군락과의 유사성이 높은 것으로 나타났다. 이는 1990년대에는 지형조건 등 환경요인에 따른 군락간 수관층의 뚜렷한 경계가 나타나지 않음을 의미한다. 선행연구결과로부터 약 30여 년이 흐른, 최근 방태산 식생구조는 지형적 조건에 따라 다소 뚜렷한 교목성 수종의 수관층이 발달한 것으로 연구결과를 도출하였으며, 이를 MRPP-test로 각 군집간 종조성의 이질성을 검증하였다. 또한 선행연구에서는 북사면에 한정한 조사였지만 본 연구결과에서는 모든 사면을 고려한 분석하였으므로 뚜렷한 식생구조로 분류된 것으로 판단된다.
패치형 모자이크 형태는 특정, 식생 발달 단계를 구별한다. 또한 식생 구조의 동적 모델을 의미하기도 한다. 하지만 이런 패치 모자이크 모델은 식생 구조를 해석하는데 있어 너무 단순하며, 양적 공간 분석이 용이하지 않다는 비판을 받아왔다(Gratzer et al., 2004; Podlaski, 2008). 그럼에도 불구하고 패치 모자이크 개념은 여러 생태학자들에 의하여 시사되는 바가 많으며(Diaci et al., 2011; Heiri et al., 2012; Huber, 2011; Kucbel et al., 2012), 숲의 구조와 역학을 설명하는데 널리 사용되고 있으며, 생물다양성 평가를 위한 정책적 프레임 워크로 사용된다(Boncina, 2000; Winter and Brambach, 2011). 패치형 모자이크 식생 구조는 안정된 구조로 보여지지만 사실상 생태계는 비평형 모델(non-equilibrium model)이라고 불리는 이론에서 벗어날 수 없으며(Gillson, 2004), 안정적인 식생 구조는 군집 간 상대적 교란의 정도를 의미한다.
이에 따라 상대적으로 동적인 패치형태를 가지는 군집은 C3와 C4로 판단된다. 그 이유는 C3의 경우 분비나무 군집으로 국내 아고산 침엽수를 대표하는 식생이다(Park et al., 2022). 본 서식지 환경은 암석노출도가 많으며, 경사도가 높아 토양발달이 불량하며 해발고도가 높아 풍해의 위험이 높아 빈번한 숲틈(Forest gap)발생이 일어나는 지역이다(Germino et al., 2002; Han et al., 2012; Hasegawa and Mori, 2007; Kim et al., 2019; Kong, 2002). 이로 인하여 분비나무의 strong association 상관관계 분석에서 미역줄나무와 시닥나무, 미역줄나무와 같은 수종과의 연관성이 있는 것으로 분석되었는데, 미역줄나무는 국내 2차림 내 광량이 노출된 교란지에서 빈번하게 번성하는 만경류 식물이다(Park et al., 2016; Park et al., 2024). 또한 국내 아고산 침엽수림 내 시닥나무, 부게꽃나무, 청시닥나무 등 Acer 속이 널리 분포되어 있는데, 이들은 내한성과 내음성 식물로 전나무속(Abies)과의 수관 경쟁이 유력한 목본식물이다(Park et al., 2021; Park et al., 2024). 또한 Acer 속은 교란 후, 근주맹아를 통한 갱신이 활발한 수종으로(Park et al., 2021), 향후 실생묘로 번식하는 분비나무림과의 임분 밀도 경쟁에서 도태될 가능성이 있는 곳으로 방태산 능선부를 중심으로 멸종위기 아고산 침엽수의 보전을 위한 지속적인 모니터링이 요구된다. C3군집의 교란의 상대적 빈번함은 지표종 분석에서도 나타나는데, 특정 군집 내 지표종이 많이 선정된 것은 해당 식생의 생물다양성이 다양함을 의미하기도 하나, 분비나무 군집을 중심으로한 외부 식물의 유입기회가 높음을 의미하기도 한다.
조릿대 군집인 C4는 국내 넓은 식생분포 내 산불 등 대규모 교란 이후 발생한 산지가 대부분이며, 전형적인 2차림 형태의 식생 구조를 나타낸다. 조릿대는 대나무류에 속하여 일제히 개화하고 종자발아 후, 고사하는 게릴라성 생활사를 나타내어, 식생 동태가 매우 활발한 군집이다(Park and Choi, 2017). 조릿대 군집은 천이 초기 타감물질을 분비함으로써, 타 종의 발아를 억제시켜 군집 내 종다양도를 낮춘다(Hiura et al., 1996; Kim et al., 1998; Kunisaki, 2004; Peters et al., 1992). 그러나 천이 후기에 들어와서는 오히려 토양 발달을 증가시키며 강한 맹아력을 지닌 내음성 수종의 생육에 긍정적인 기여를 하기도 하여(Cha and Chun, 2002; Yamamoto et al., 1995) 향후 조릿대 군집은 내음성 수종인 Acer 속 또는 아고산지대에 주로 분포하는 사스래나무 등 Betula 속의 식생 구조로 변화할 것으로 판단된다.
한편, 패치형 모자이크 식생 구조는 군집과 군집사이 생태적 점이지대(ecotone)라는 전이구역을 형성한다(Cadenasso et al., 2003). 이는 각 패치의 둘레를 나타내는 가장자리(edge)를 나타낸다. 생태적 점이지대는 다양한 조건과 서식지 환경을 조성하여(Turner. 1989; Urban et al., 1987), 본 연구대상지인 방태산 일대의 생물다양성이 풍부하고 식생 동태가 활발한 지역이 곳곳에 존재할 것으로 판단된다. 가장자리에서는 물질순환, 동식물의 흐름을 통하여 각각의 패치를 기능적으로 연결하기 때문이다. 가장자리에서는 식생 동태가 활발하고, 교란이 빈번하여 호광성 식물종이 번성하고 건조 스트레스를 견디는 능력이 있다(Cadenasso et al., 2003). 생태적 전이지역 내 다양한 서식처를 제공하여 동물 종의 생물다양성의 변화에 영향을 주기도 한다. 종합하면 생태적 전이지역은 식생 동태가 매우 활발하며, 다양한 생물다양성을 증진시킬 수 있으며, 패치 사이의 생태계 네트워크의 역할을 한다. 그러나 국내 산림생태계 내 패치사이의 생태적 점이지대에 대한 식생 변화 및 발달 연구가 미흡하여 본 연구대상지가 좋은 사례연구지가 될 것으로 판단된다.
보호구역의 지정은 생물다양성을 보전하는 매우 중요한 정책옵션이라고 할 수 있다. 그러나 최근 쿤밍-몬트리올 글로벌 프레임 워크에서는 기타 효과적 지역기반 보전수단(Other Effective Area-Based Conservation Measures, OECMs)을 통하여 생태계 보전을 위하여 보호구역 뿐만아니라, 다방면의 방법으로 생물다양성 유지 및 증진을 수행할 수 있다(CBD, 2024; Hwang et al., 2023). 따라서, 생태계를 보전하는데 있어, 생태학 연구자들은 특정 서식지를 대상으로 다각적인 접근으로 보전 정책옵션 제시를 위한 과학적 근거자료 마련이 필요하다.
적 요
본 연구는 WDPA에 속해있으며, 국내 산림유전자원보호구역인 방태산의 산림 식생 구조를 분석하여, 연구대상지의 보전전략 및 효율적인 관리를 위한 기초자료를 마련하는 데 있다. 식생군집은 크게 4개 군집으로 분류되었으며, 당단풍나무-고로쇠나무 군집(C1), 신갈나무 군집(C2), 분비나무 군집(C3), 그리고 조릿대-당단풍나무 군집(C4)로 분류되었다. 군집 간 지형 환경 인자는 TPI를 제외한 인자에서 통계적 유의성이 인정되어, 각 군집이 고유한 지형적 특성을 지니고 있음을 확인하였다. 종조성 분석 결과, 당단풍나무와 고로쇠나무와 같은 Acer 속이 계곡부에 생육하고 있었으며, 아고산대 식물인 분비나무와 사스래나무도 고루 분포하고 있다. 방태산 산림유전자원보호구역 일대는 아고산 및 온대림 식생이 혼합된 형태로 나타났다. 이에 따라 지형적 특성과 같은 복합적인 환경요인에 따라 군집 분류가 뚜렷하게 나타나는 것으로 보아 패치 모자이크 형태의 식생 분포를 나타내고 있었다. 방태산 산림유전자원보호구역의 식생 패치 사이의 생태적 점이지대가 넓게 분포하고 있을 것으로 판단되며, 이는 방태산 산림유전자원보호구역 내 풍부한 생물다양성을 보유하며, 원활한 물질순환이 이루어져 건강한 산림생태계를 유지하고 있음을 의미한다. 따라서 향후 연속적인 모니터링과 식생 변화 관찰을 통한 보전전략의 마련이 필요함을 시사한다.
