

Introduction
Materials and Methods
Plant materials
Primary culture of two Allium species
Callus induction in Allium explants
Culture conditions and data analysis
Results
Callus induction
Root tip responses
Bulb meristem responses
Discussion
PGR effects on callus induction
Explant-dependent organogenic responses
Contamination in bulb meristem cultures
Practical use of root tip explants
Introduction
Allium species (Amaryllidaceae) are highly diverse: they exhibit considerable variation in floral morphology, color, and inflorescence characteristics (Fritsch, 2015). Although edible Allium species are more well known, approximately 250 Allium species, including A. caeruleum, A. caesium, A. giganteum, A. schubertii, and A. sphaerocephalon, have been reported to have commercial value as ornamental plants (Fritsch, 2015).
Among the native Korean Allium species, A. dumebuchum H.J.Choi, which is endemic to Korea (Jang et al., 2021), and A. thunbergii G.Don are of particular interest as potential ornamental resources due to their distinctive inflorescence characteristics and floral traits, especially perianth color. A. dumebuchum exhibits a light-purple perianth color, whereas A. thunbergii bears a red-to-purple perianth (Jang et al., 2024). In addition to their ornamental value, these species are important native genetic resources for conservation and sustainable use. Conventional propagation of native ornamental geophytes can be limited by the low multiplication rate of bulb-derived materials, seasonal growth cycles, and the need to avoid excessive collection from natural populations. In this context, in vitro propagation may offer a practical means of producing plant materials while supporting ex situ conservation of native Allium species.
Plant tissue culture allows plant cells, tissues, or organs to be cultured aseptically under controlled conditions and is widely used for propagation and regeneration (Pan et al., 2024). Callus culture, in particular, involves the induction of dedifferentiated tissue from explants, which can subsequently redifferentiate into shoots, roots, or somatic embryos depending on genotype, explant type, and plant growth regulator (PGR) conditions. This technique has been widely used in studies of plant cell differentiation and development, mass propagation, germplasm conservation, breeding, and metabolite production (Bai et al., 2024; Mosa et al., 2025; Seo et al., 2025).
Callus induction and subsequent morphogenic responses are largely determined by the composition of the culture medium, especially the type and concentration of PGR. Auxins and cytokinins are the principal classes of PGR used to regulate dedifferentiation and organogenesis in vitro (Jalilian et al., 2025). Auxins such as 2,4-dichlorophenoxyacetic acid (2,4-D) and 1-naphthaleneacetic acid (NAA) are commonly used for callus induction, whereas cytokinins such as 6-benzylaminopurine (BAP) and thidiazuron (TDZ) are often applied to promote shoot organogenesis or related morphogenic responses. However, the response to these PGR can vary depending on plant species, explant type, and culture conditions.
These hormonal effects have been reported in various herbaceous and woody plant species, such as Camellia sinensis (0.2 ㎎·L-1 NAA + 3.0 ㎎·L-1 BAP) (Zhang et al., 2025), Iris pseudacorus (2.0 ㎎·L-1 2,4-D + 0.2 ㎎·L-1 kinetin) (Li et al., 2025), Peucedanum praeruptorum (2.0 ㎎·L-1 2,4-D + 0.5 ㎎·L-1 BAP) (Pan et al., 2024), Stachys inflata (0.5 ㎎·L-12,4-D + 0.1 ㎎·L-1 BAP) (Jalilian et al., 2025), and Tephrosia apollinea (2.0 ㎎·L-1 2,4-D + 0.25 ㎎·L-1 BAP) (Mosa et al., 2025). In Allium, callus induction has been investigated to a limited extent in edible species such as garlic (Luciani et al., 2006), leek (Buiteveld et al., 1993), onion (Zheng et al., 1998), and Persian shallot (Farhadi et al., 2017).
As callus induction and redifferentiation responses in Allium may vary considerably depending on the species or genotype (Barandiaran et al., 1999; Mostafa et al., 2020), species-specific callus culture systems for the micropropagation of Allium species need to be established. However, the PGR requirements for callus induction in ornamental Allium species remain poorly elucidated. Hence, this study was conducted to determine the conditions that promote callus induction in bulb meristem and root tip explants from two Allium species native to Korea: A. dumebuchum and A. thunbergii. The findings of this study provide useful baseline information for developing species-specific in vitro culture protocols for the propagation, conservation, and horticultural utilization of ornamental Allium species native to Korea.
Materials and Methods
Plant materials
In 2019, A. dumebuchum and A. thunbergii plant samples were collected from their natural habitats in Ulleungdo and Pyeongchang, South Korea. Subsequently, the samples were transplanted to the Korea National Arboretum in Yangpyeong. Seeds from the transplanted individuals were collected and stored at 4°C until their further use in primary culture experiments conducted in 2020.
Primary culture of two Allium species
Primary cultures for the in vitro introduction of A. dumebuchum and A. thunbergii were established using bulb meristems and seeds. For bulb-derived cultures, soil and other debris were removed from bulb samples under running tap water. Afterwards, the bulbs were disinfected in 75% ethanol for 1 min, surface-sterilized in 2% sodium hypochlorite (NaClO) for 15 min with shaking, and rinsed at least three times with sterile water. After sterilization, fully developed bulb scales were removed, and meristems approximately 2–3 ㎜ in size were aseptically excised and inoculated onto a culture medium.
For seed-derived cultures, seeds were washed thoroughly with tap water, disinfected in 75% ethanol for 5 min, surface-sterilized in 2% NaClO for 15 min, and rinsed several times with sterile water. The sterilized seeds were germinated in vitro on a Murashige and Skoog (MS) basal medium, consisting of MS (4.4 g·L-1), sucrose (30 g·L-1), and agar (6 g·L-1). The pH of the basal medium was adjusted to 5.7–5.8 before autoclaving. After germination, root tip segments approximately 2–3 ㎜ in length were excised and used as explants.
Callus induction in Allium explants
The bulb meristem and root tip explants established in vitro were cultured on a PGR-free MS medium (control) or an MS medium supplemented with 0.1, 0.5, or 1.0 ㎎·L-1 NAA (Sigma-Aldrich, St. Louis, MO, USA), 2,4-D (Sigma-Aldrich), BAP (Duchefa Biochemie B.V., Haarlem, The Netherlands), or TDZ (Duchefa Biochemie B.V.). Ten explants were placed per Petri dish. Callus induction and redifferentiation were evaluated after 3 months of culture. Explants exhibiting initial contamination were included in the assessment as long as callus formation, and redifferentiation was confirmed. The numbers of bulblets, shoots, and roots were recorded. Callus induction frequency was calculated for each Petri dish as follows:
Culture conditions and data analysis
Unless otherwise stated, all cultures were maintained at 24°C under a photosynthetic photon flux density of 80 µmol·m-2·s-1, a 16:8 h light:dark photoperiod, and 50% relative humidity. The experiment followed a randomized complete block design with three replicate Petri dishes per treatment. Because replicate-based calculations were available for callus induction frequency, statistical analysis was performed for this variable. The callus induction frequency values from the three replicate Petri dishes were used to calculate the mean and standard error for each treatment. Differences among treatments were analyzed by performing one-way analysis of variance, followed by Duncan’s multiple range test at P < 0.05. All statistical analyses were conducted using SAS software (version 9.4; SAS Institute Inc., Cary, NC, USA).
Results
Callus induction
Callus induction responses differed between A. dumebuchum and A. thunbergii and varied according to explant type and PGR treatment. Callus was induced from root tip explants of both species. In A. dumebuchum, root tip explants cultured on auxin-containing media showed relatively high callus induction frequencies. Among the auxin-containing treatments, the highest frequency, 90.0%, was observed on medium supplemented with 1.0 ㎎·L-1 NAA (Fig. 1A). In A. thunbergii, root tip explants showed the highest callus induction frequency, 100.0%, on medium supplemented with 0.1 ㎎·L-1 2,4-D (Fig. 1B). Callus was also induced from bulb meristem explants. In A. dumebuchum, bulb meristem explants formed callus under all treatments except 2,4-D, and BAP induced callus formation at a frequency of 100.0% regardless of concentration (Fig. 1C). In A. thunbergii, bulb meristem explants showed a more restricted response, with callus induction observed only on media supplemented with 0.5 ㎎·L-1 BAP and 0.5–1.0 ㎎·L-1 TDZ (Fig. 1D). Among the cytokinin treatments, only 1.0 ㎎·L-1 BAP induced callus formation in root tip explants from both species, with callus induction frequencies of 60.0% in A. dumebuchum and 30.0% in A. thunbergii. In bulb meristems, BAP and TDZ were more frequently associated with callus formation and organogenic responses. Representative images of callus induction and related morphogenic responses in root tip and bulb meristem explants are shown in Fig. 2. The selected panels highlight the contrasting responses of the two explant types under PGR treatments that induced callus formation in each species.
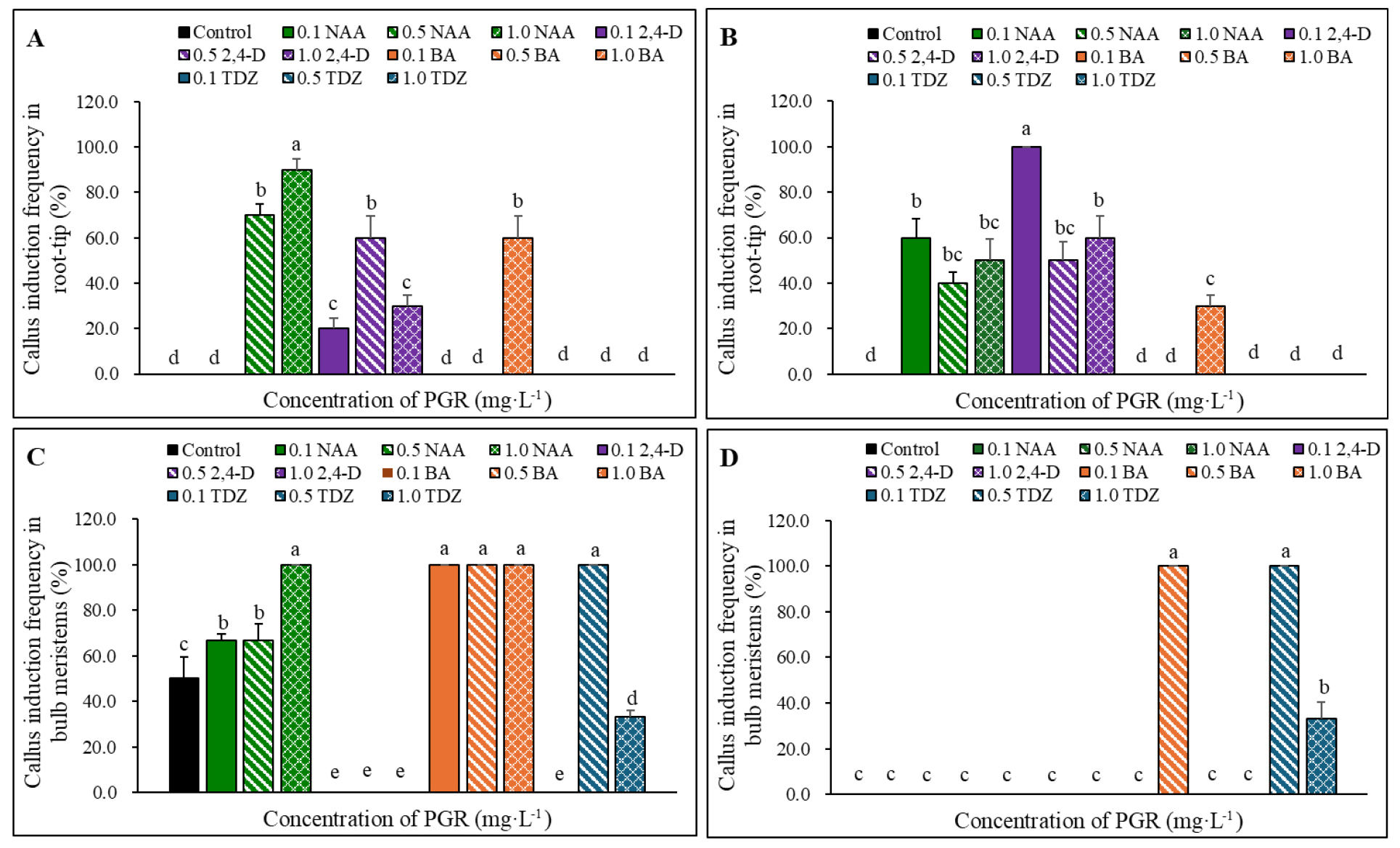
Fig. 1.
Effects of plant growth regulator (PGR) on callus induction in root tip and bulb meristem explants from Allium dumebuchum and A. thunbergii. Callus induction frequency was evaluated after 3 months of culture in root tip explants from A. dumebuchum (A) and A. thunbergii (B), and bulb meristem explants from A. dumebuchum (C) and A. thunbergii (D) cultured on a PGR-free MS medium (control) or an MS medium supplemented with 0.1, 0.5, or 1.0 ㎎·L-1 NAA, 2,4-D, BAP, or TDZ. Lowercase letters above the bars indicate significant differences among treatments within each panel at p < 0.05 according to Duncan’s multiple range test. Control, PGR-free MS medium; NAA, 1-naphthaleneacetic acid; 2,4-D, 2,4-dichlorophenoxyacetic acid; BAP, 6-benzylaminopurine; TDZ, thidiazuron.
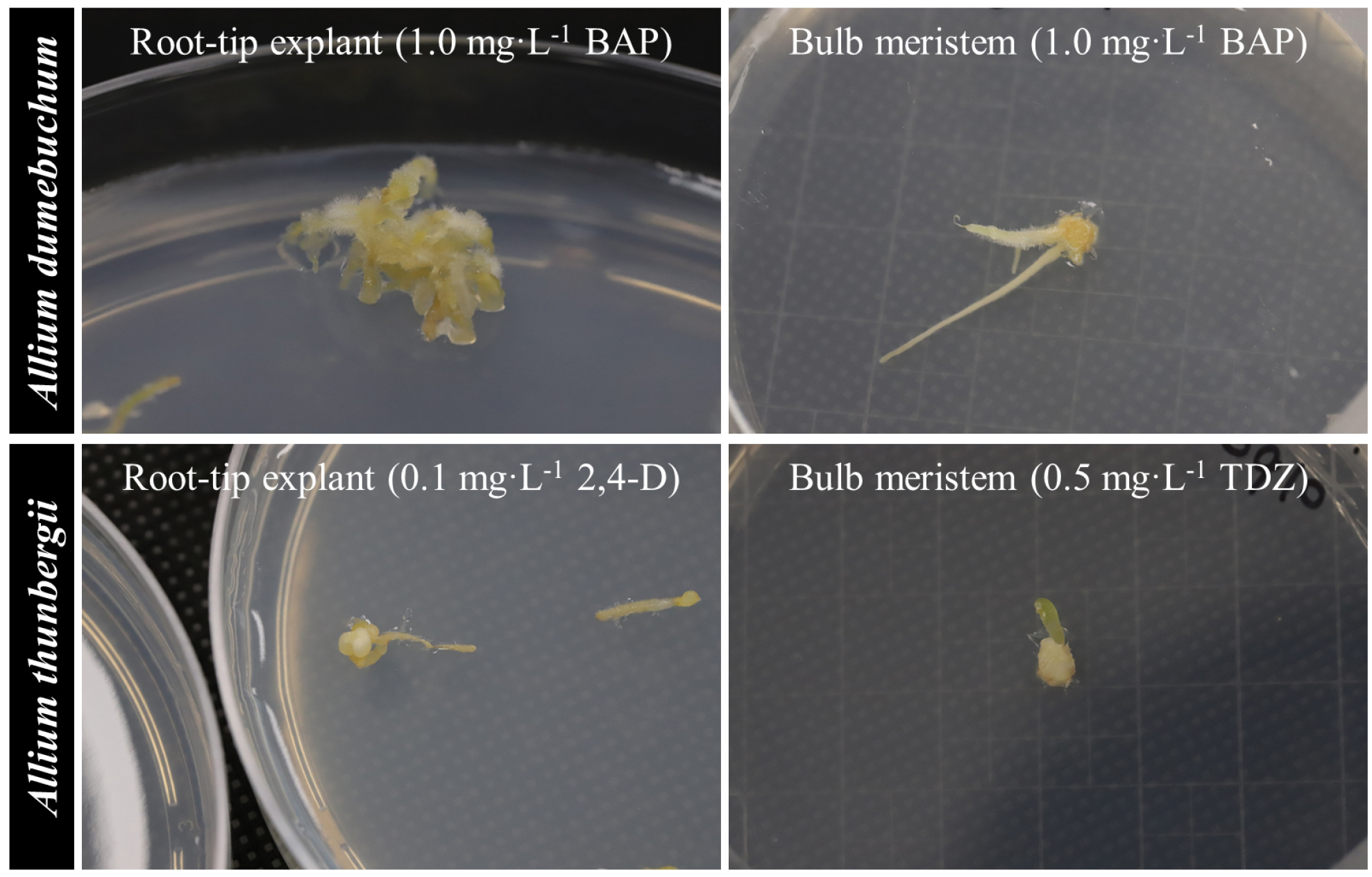
Fig. 2.
Representative images of callus induction and related morphogenic responses in root tip and bulb meristem explants from Allium dumebuchum and A. thunbergii cultured for 3 months on media supplemented with different plant growth regulators. (A) A root tip explant from A. dumebuchum cultured on 1.0 ㎎·L-1 BAP. (B) A bulb meristem explant from A. dumebuchum cultured on 1.0 ㎎·L-1 BAP. (C) A root tip explant from A. thunbergii cultured on 0.1 ㎎·L-1 2,4-D. (D) A bulb meristem explant from A. thunbergii cultured on 0.5 ㎎·L-1 TDZ. 2,4-D, 2,4-dichlorophenoxyacetic acid; BAP, 6-benzylaminopurine; TDZ, thidiazuron.
Root tip responses
Regeneration-related responses were limited in root tip explants and manifested mainly as root formation rather than shoot or bulblet development (Tables 1 and 2). In A. dumebuchum, root tip explants did not produce bulblets or shoots under any PGR treatment. Nonetheless, the culture medium supplemented with 1.0 ㎎·L-1 NAA produced the highest number of roots, 3.8, followed by 0.5 ㎎·L-1 NAA, 1.9 roots, and the control, 1.1 roots (Table 1). In A. thunbergii, root tip explants also showed limited responses. The culture medium supplemented with 1.0 ㎎·L-1 NAA produced the highest number of roots, 2.3. On the other hand, 0.1 ㎎·L-1 TDZ induced low levels of bulblet, 0.2, shoot, 0.6, and root, 0.4, formation, whereas 1.0 ㎎·L-1 TDZ induced the formation of only 0.2 root (Table 2).
Table 1.
Regenerative responses of Allium dumebuchum explant types to plant growth regulator (PGR) treatments.
| Explant types | PGR treatmentsz (㎎·L-1) | No. of bulblets | No. of shoots | No. of roots |
| Root tip | Control | 0 | 0 | 1.1 |
| 0.1 NAA | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 1.9 | |
| 1.0 | 0 | 0 | 3.8 | |
| 0.1 2,4-D | 0 | 0 | 0.4 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 BAP | 0 | 0 | 0.2 | |
| 0.5 | 0 | 0 | 0.2 | |
| 1.0 | 0 | 0 | 0.1 | |
| 0.1 TDZ | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| Bulb meristems | Control | 0 | 0.5 | 0.5 |
| 0.1 NAA | 0 | 0.7 | 0.7 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 2,4-D | 0 | 0 | 0 | |
| 0.5 | 0 | 1.0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 BAP | 0 | 0 | 2.5 | |
| 0.5 | 0 | 6.3 | 0 | |
| 1.0 | 0 | 0 | 1.0 | |
| 0.1 TDZ | 0 | 2.0 | 1.5 | |
| 0.5 | 0 | 0.3 | 0.3 | |
| 1.0 | 0 | 0 | 0 |
Table 2.
Regenerative responses of Allium thunbergii explant types to plant growth regulator (PGR) treatments.
| Explant types | PGR treatmentsz (㎎·L-1) | No. of bulblets | No. of shoots | No. of roots |
| Root tip | Control | 0 | 0 | 0 |
| 0.1 NAA | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 2.3 | |
| 0.1 2,4-D | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 0.1 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 BAP | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 TDZ | 0.2 | 0.6 | 0.4 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0.2 | |
| Bulb meristems | Control | 0 | 6.8 | 1.5 |
| 0.1 NAA | 0 | 0 | 0 | |
| 0.5 | 0 | 0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 2,4-D | 0 | 0 | 0 | |
| 0.5 | 0 | 1.0 | 0 | |
| 1.0 | 0 | 0 | 0 | |
| 0.1 BAP | 0 | 1.5 | 1.7 | |
| 0.5 | 0 | 2.3 | 1.3 | |
| 1.0 | 0 | 2.3 | 4.0 | |
| 0.1 TDZ | 0 | 2.0 | 1.5 | |
| 0.5 | 0 | 0.3 | 0.3 | |
| 1.0 | 0 | 0 | 0 |
Bulb meristem responses
In contrast, bulb meristem explants exhibited more pronounced shoot and root formation, although these responses were inconsistent across treatments (Tables 1 and 2). In A. dumebuchum, 0.5 ㎎·L-1 BAP produced the highest number of shoots, 6.3, whereas 0.1 ㎎·L-1 BAP produced the highest number of roots, 2.5. In addition, 0.1 ㎎·L-1 TDZ induced the formation of 2.0 shoots and 1.5 roots, and 0.5 ㎎·L-1 2,4-D induced the formation of 1.0 shoot (Table 1). In A. thunbergii, the control produced the highest mean number of shoots, 6.8, whereas 1.0 ㎎·L-1 BAP produced the highest mean number of roots, 4.0. BAP at 0.1–0.5 ㎎·L-1 induced the formation of 1.5–2.3 shoots and 1.3–1.7 roots, and 0.1 ㎎·L-1 TDZ induced the formation of 2.0 shoots and 1.5 roots (Table 2).
Discussion
PGR effects on callus induction
Callus induction from root tip explants in both species agrees with previous reports that callus can be readily formed in the meristematic zone of Allium root tips (Robledo-Paz et al., 2000; Tubić et al., 2014). In the present study, the effective PGR treatment differed between the two species. For A. dumebuchum, 1.0 ㎎·L-1 NAA resulted in the highest callus induction frequency from root tip explants, whereas 0.1 ㎎·L-1 2,4-D was most effective for A. thunbergii. These results are consistent with previous studies showing that auxins, particularly 2,4-D, are effective for callus induction in Allium species (Buiteveld et al., 1993; Jalilian et al., 2025; Zheng et al., 1998). However, the different responses to NAA and 2,4-D indicate that callus induction conditions should be optimized separately for each species, even within the same genus. Under the present culture conditions, seed-derived root tip explants were more suitable than bulb meristems for establishing aseptic callus cultures of A. dumebuchum and A. thunbergii.
Explant-dependent organogenic responses
Root tip explants were mainly associated with callus induction and root formation, whereas shoot or bulblet development from this explant type was limited. In A. dumebuchum, root tip explants did not produce shoots or bulblets under any PGR treatment. In A. thunbergii, only low levels of bulblet and shoot formation were observed under TDZ treatment. These results suggest that root tip explants can serve as a useful starting material for callus induction, but their capacity for shoot regeneration was limited under the present conditions. Further refinement of PGR combinations or culture procedures will be needed to improve shoot or bulblet formation from root tip-derived callus.
Contamination in bulb meristem cultures
Contamination represents an important limitation in interpreting the bulb meristem data. The bulb-derived cultures were contaminated during primary culture, probably because belowground organs, such as bulbs and rhizomes, generally carry relatively high microbial loads (Hol and van der Linde, 1992). Although surface sterilization with 2% NaClO for 15 min, which is commonly used for explant disinfection, was applied in this study, it was insufficient to control contamination. Therefore, improved disinfection procedures or antimicrobial supplementation may be required for a more rigorous evaluation of bulb-derived explants and for the propagation of disease-free plantlets (Leelavathy and Sankar, 2016). The apparent superiority of seed-derived root tip explants should thus be interpreted mainly in terms of practical aseptic culture establishment under the present conditions rather than as definitive evidence that bulb meristems have lower intrinsic morphogenic potential.
Practical use of root tip explants
From a practical perspective, the present results support the use of seed-derived root tip explants as a more feasible starting material for callus induction in A. dumebuchum and A. thunbergii under the experimental conditions in this study. In particular, 1.0 ㎎·L-1 NAA was the most effective PGR treatment for root tip-derived callus induction in A. dumebuchum, whereas 0.1 ㎎·L-1 2,4-D was the most effective in A. thunbergii. However, the organogenic responses of A. dumebuchum and A. thunbergii were limited and inconsistent. These preliminary results may serve as a reference for establishing a more robust plant regeneration system from root tip explants. Overall, the findings provide baseline information for future studies aimed at optimizing callus induction and more rigorously evaluating regeneration potential in other wild or ornamental Allium species.
